Стійкість цукрових буряків до посухи: фізіологічні та молекулярні аспекти (частина 2)

За матеріалами розділу на тему «Стійкість цукрових буряків до посухи: фізіологічні та молекулярні аспекти» («Sugar beet tolerance to drought: physiological and molecular aspects») книги «Рослина, абіотичний стрес та реакції на зміни клімату» («Plant, abiotic stress and responses to climate change») під редакцією Віолети Анжелькович, опублікованої 23 травня 2018 року.
Результати та обговорення
Як було зазначено раніше (Таблиця 1 та 2), кліматичні умови регіону вказують на необхідність проведення досліджень, які можуть покращити вибір генотипів, більш стійких до посухи.
Експеримент у напів-контрольованих умовах у теплиці
Класифікація генотипів цукрових буряків на основі фізіологічних тестів у напів-контрольованих умовах
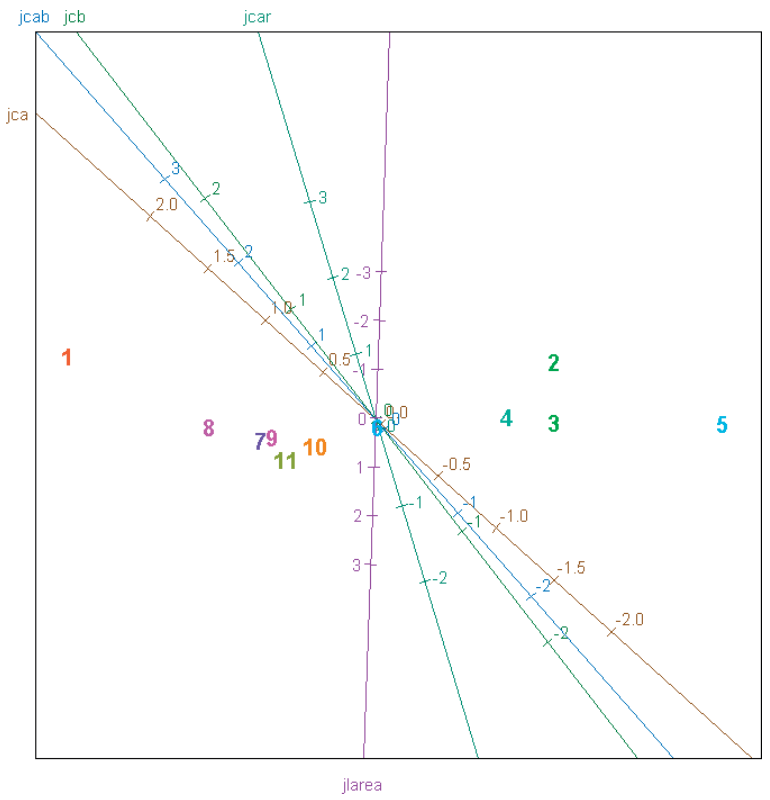
У напів-контрольованих умовах генотипи цукрових буряків виявляли різні реакції на 5-денний дефіцит води. Як і очікувалося, зниження тургору спостерігалось у всіх генотипах. Кількість листків значно відрізнялася, в залежності від обробок та відповідних засобів захисту. Концентрації фотосинтетичних пігментів та площа листя також змінювалися між генотипами, а стандартний нормальний розподіл був відсутнім. Отже, отримані результати були піддані перетворенню даних Джонсона, що виявилося дуже ефективним та дозволило визначити відмінності концентрацій фотосинтетичних пігментів у різних генотипах (Графік 1).
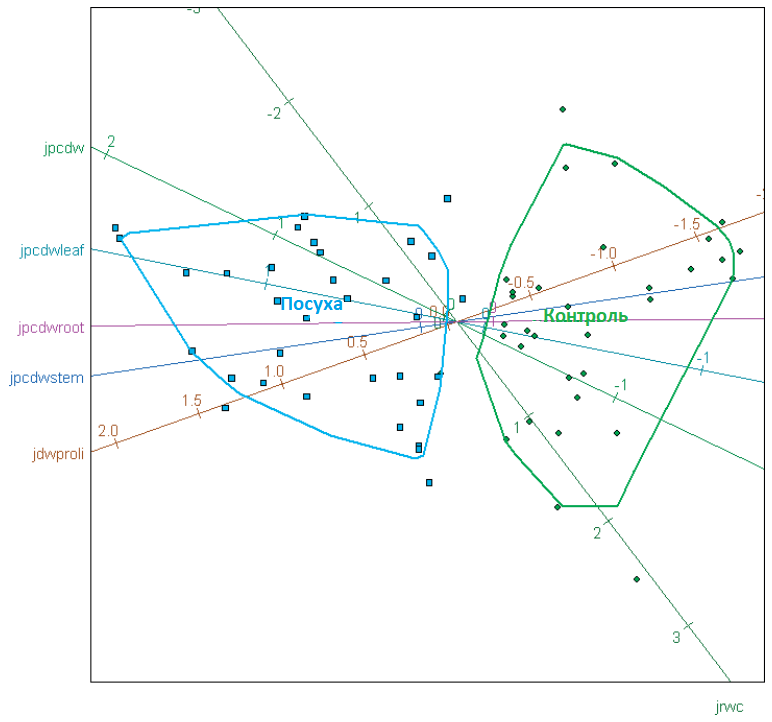
Сецесія водопостачання спричинила втрати води з рослинних тканин як у чутливих, так і в стійких генотипах. У зв'язку з цим, генотипи цукрових буряків можна розділити на основі досліджуваних параметрів та методів обробок (Графік 2).
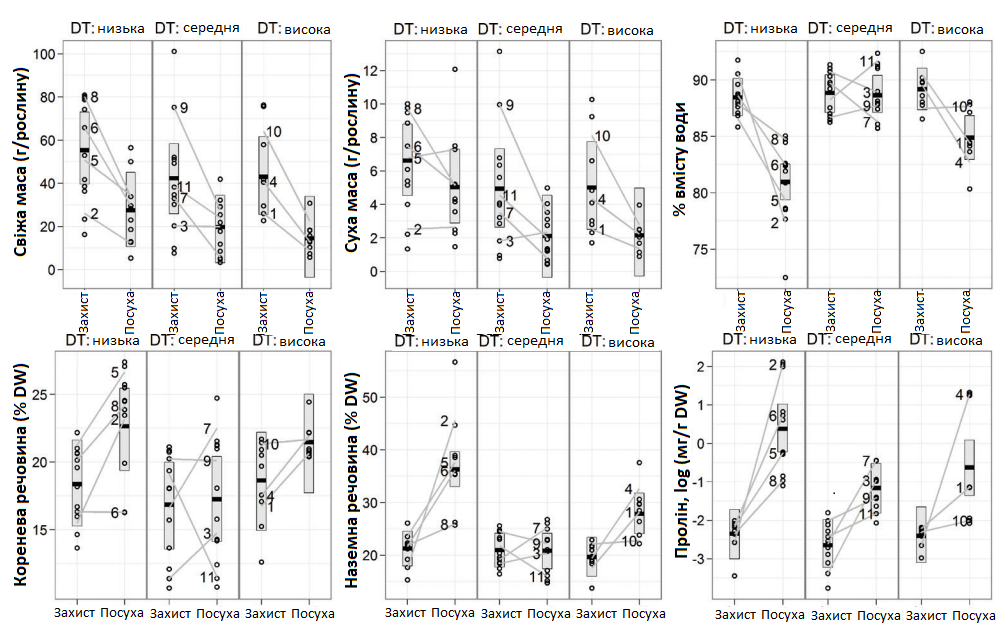
Результати, отримані в напів-контрольованих умовах (експеримент 1), порівнювалися з попередніми польовими спостереженнями (Графік 3). У результаті впливу дефіциту води, у всіх генотипах (окрім генотипів 9 та 11) збільшилася концентрація проліну, а також % сухої речовини (DM). Зміни в рамках захисних обробок, що стосуються сухої маси, були менш вираженими, а ніж зміни, що стосуються % сухої речовини (DM) та вмісту води в корені, стеблі та листі (RWС). Рослини, що перебували в стресових умовах, мали в середньому на три листка менше, на 4% більший % сухої речовини (DM) та в сім разів більший вміст проліну.
Графік 1. Розподіл генотипів на основі концентрації пігменту та площі листя для змінних, нормалізованих згідно з трансформацією даних Джонсона (jlarea — площа листя; jcar — каротиноїди; jca — хлорофіл a; jcb — хлорофіл b; jcab — хлорофіл a + b).
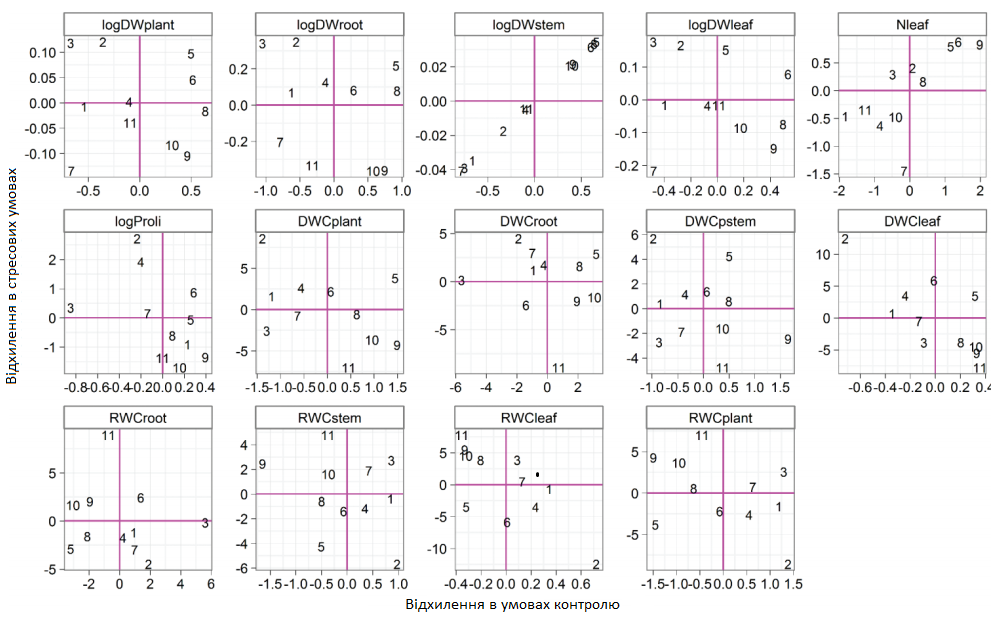
Взаємозв'язок між двома впливами на вимірювані ознаки визначався за допомогою моделі змішаного типу (Графік 4). Перехресні рожеві лінії на діаграмах представляють середній вплив на генотипи під захистом (вісь x) та вплив стресу (вісь y). Існує майже ідеальна негативна кореляція між нестресовим значенням та реакцією на стрес для сухої речовини кореня та аналогічна, але слабша кореляція для кількості листків. Генотипи, що демонструють позитивні результати (вісь y), відчувають менший середнього рівня вплив (більш стійкі до) водного стресу для відповідної ознаки, і навпаки. Генотипи, що показують позитивні результати для обох впливів, мають кращі результати за відсутності стресу і відчувають меншими середнього рівня вплив (більш стійкі до) стресу за відповідною ознакою, і навпаки.
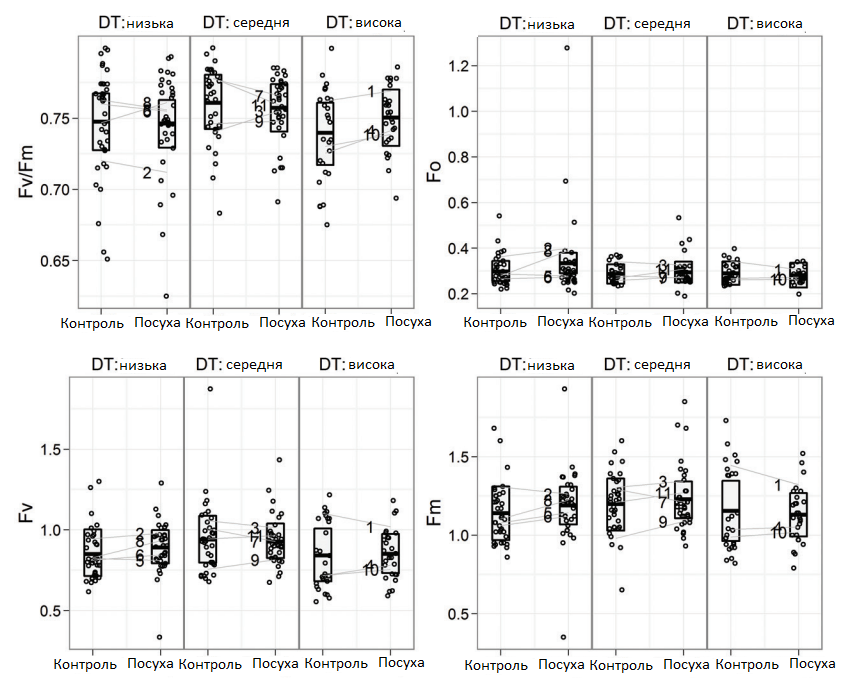
Зазвичай результати флуоресценції хлорофілу свідчать про високу чутливість до впливу екологічних факторів. Тому їх часто використовують як показник функціонування фотосистеми II.
За даними Fv/Fm, генотипи цукрових буряків порівнювалися на основі фотосинтетичних характеристик. Дефіцит води не викликав значних змін у показниках цвітіння (Графік 5).
Графік 2. Розподіл генотипів цукрових буряків на основі експериментів у теплиці з виділенням обробки (контроль, посуха) для змінної, нормалізованої згідно з трансформацією даних Джонсона (jrwc — відносний вміст води; jpcdw — суха маса; jdwproli — вільний пролін; jpcdwleaf — суха речовина в листі; jpcdwstem — суха речовина в стеблі; jpcdwroot — суха речовина в корені).
Вплив посухи спостерігався у випадку Fv і Fm, але не для співвідношення Fv/Fm, де було виявлено найбільші відмінності між генотипами. Крім того, інтервали взаємодії між стресом і генотипом вказують на стрес, який спричинив відмінності, подібні для всіх генотипів. Вплив дефіциту води на флуоресценцію може бути пов'язаний зі стійкістю рослин до дефіциту води у польових умовах (Графік 5).
У польових умовах розвиток рослин може сповільнюватися різними факторами. На нього можуть впливати взаємодія посухи з іншими екологічними стресами, опадами та температурою, а також із різними мікроорганізмами. З іншого боку, напів-контрольовані умови можуть виключити лише втручання інших факторів у розвиток рослин. Тому необхідно порівнювати результати, отримані в тепличних умовах, із результатами, отриманими в польових умовах.
Експеримент по експлантації в лабораторних умовах
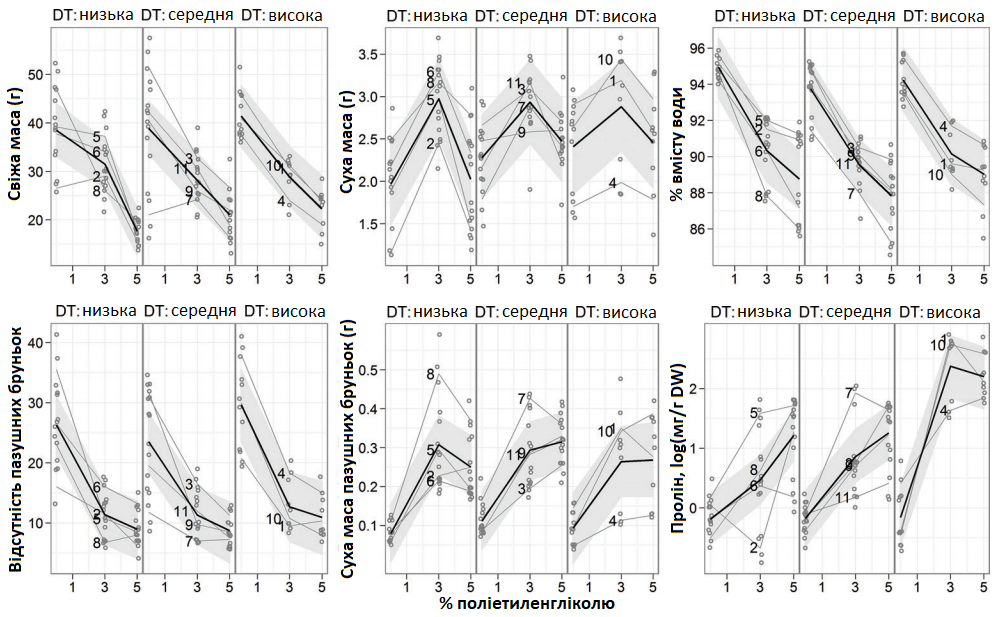
Підвищена концентрація поліетиленгліколю (PEG) зменшила ріст бічних пагонів, порівняно з захистом (Графік 6).
Графік 3. Вплив стресу, викликаного посухою, на параметри росту та виробництво проліну в генотипах цукрових буряків, вирощених у теплиці (1-11), з трьома класами візуальної оцінки стійкості до посухи (DT) в польових умовах. Отримані значення трьохкратної повторюваності (кола, по 10 рослин у кожному), середні позиції генотипів (пронумеровані сірі лінії) та класові середні значення із 95% довірчим діапазоном (горизонталі). Стрес від посухи був викликаний припиненням поливу на дослідних ділянках, спостереження проводилися через 5 днів.
Кількість бічних пагонів може бути показником впливу різних концентрацій поліетиленгліколю (PEG), які викликають дефіцит води, на потенціал мікроклонального розмноження генотипів. Середня кількість бічних пагонів 11 досліджуваних генотипів зменшилася в 2,2 разів за наявності 3% PEG та в 2,7 разів — при 5% PEG.
Ступінь стійкості до посухи, що спостерігалася в польових умовах, відповідав стійкості, зафіксованій в експериментах, проведених у теплиці та в лабораторних умовах (Графік 6). Найбільш значимим критерієм визначення стійкості генотипу до посухи виявилася концентрація вільного проліну. Концентрація проліну помітно зростала в листі, що зазнало впливу посухи, та в бічних пагонах і позитивно корелювалася з концентрацією PEG, відповідаючи результатам інших досліджень.
Контроль PEG зменшив загальну суху масу та кількість пазушних бруньок більше ніж удвічі, тоді як наявність 3% PEG у субстраті збільшила загальну свіжу масу. Крім того, PEG викликав зменшення вмісту води в тканинах та кількості бруньок, але збільшив їх масу та % DM. Найвищі значення були зафіксовані під час контролю (0% PEG) для загальної свіжої маси, за наявності 3% PEG — для концентрації проліну та свіжої маси пазушних бруньок та при 5% PEG — для % DM. Свіжа маса рослин, вирощених при 3 та 5% PEG, зменшилася (Графік 6). Середня суха маса рослин була найвищою при наявності 3% PEG. Однак за наявності 5% PEG вона майже відповідала рівню 0% PEG. Більша мінливість сухої маси була зафіксована у чутливій до посухи групі (за даними польових спостережень), але залишилася така ж тенденція, як і в інших двох групах генотипів. Після збільшення концентрації PEG вміст води в тканинах лінійно зменшився, середнє зменшення наявності 5% PEG склало 6%, а за ним супроводжувалася низька середня різниця між групами різної стійкості та більша різниця між генотипами однієї групи (Графік 6).
Графік 4. Вплив генотипу з аналізу ознак за моделлю змішаного типу тепличного експерименту (відхилення значень в нестресових умовах на осі x, відхилення значень у стресових умовах на осі y. Генотипи ідентифікуються числами)
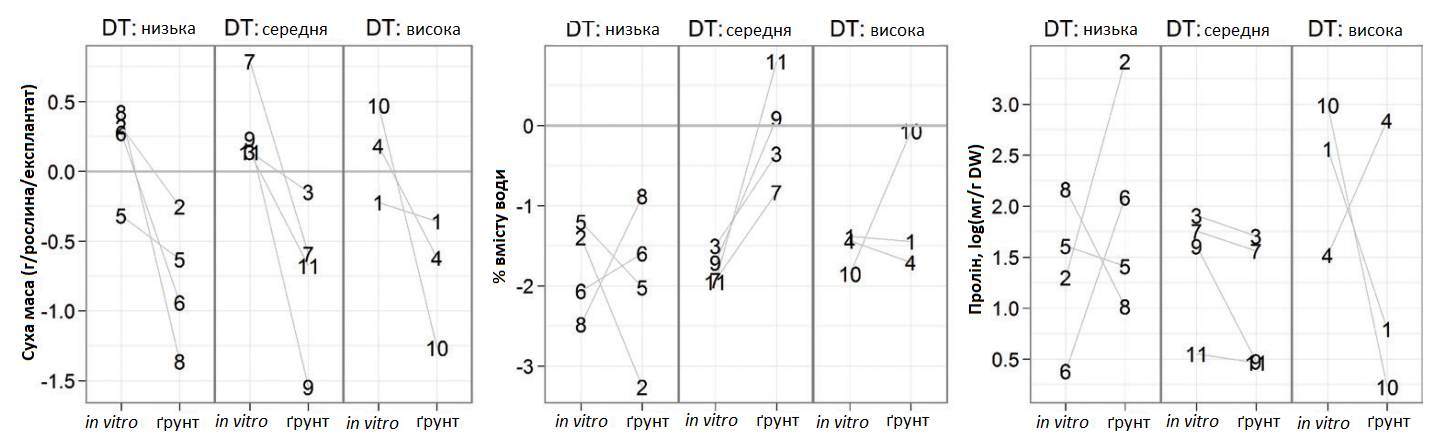
Накопичення проліну в стресових умовах збільшувалося під час захисних обробок в обох експериментах. Під час експлантації накопичення проліну збільшилося в 6 разів, а в тепличних умовах — в 16 разів.
Якщо брати до уваги стійкість генотипів у польових умовах, по відношенню до параметрів, отриманих в результаті аналізу рослин під час експлантації та в тепличному експерименті, суха речовина по відношенню до вмісту води та концентрації проліну суттєво не відрізняється серед груп генотипів різної стійкості (Графік 7).
Виявлена різниця між генотипами показує два підходи до поділу генотипів цукрових буряків за їх реакцією на водний стрес, які не можуть замінити один одного. З одного боку, вміст проліну в рослинах, вирощених в процесі експлантації, дав можливість співставити їх групування по відношенню до польових спостережень. З іншого боку, в цьому сенсі тепличний експеримент був менш ефективним (Графік 3). Основною причиною цього може бути той факт, що стрес у польових умовах не був постійним, як це було в теплиці.
Графік 5. Максимальна (Fm) та змінна (Fv) флюоресценція хлорофілу та їх співвідношення (Fv/Fm) генотипів цукрових буряків, згрупованих у відповідності до їх стійкості до посухи в польових умовах (ctrl — контроль; посуха; DT — стійкість до посухи)
Графік 6. Вплив поліетиленглюколю (PEG) на ознаки росту та концентрацію вільного проліну рослин, вирощених під час експлантації
Графік 7. Вплив водного дефіциту на суху масу, вміст води та концентрацію вільного проліну в тепличному експериме5нті та в умовах експлантації
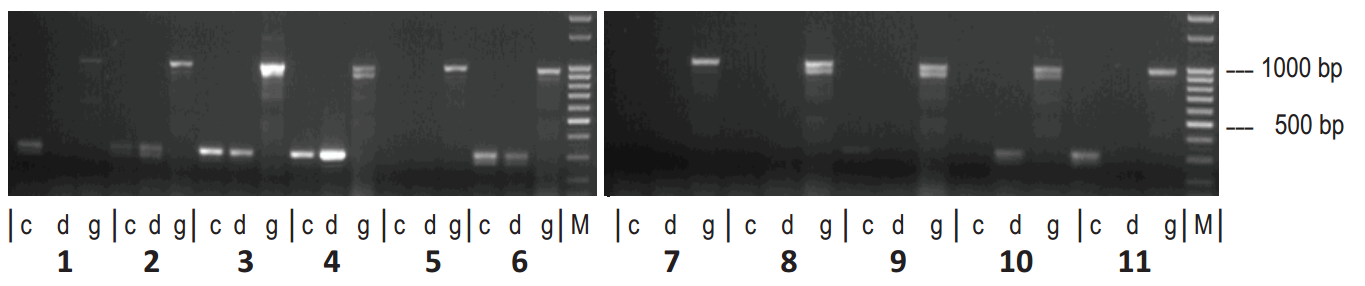
Графік 8. Схема експресії генів, що відповідає BI543243, у листі цукрових буряків (c — шаблон кДНК, що походить від контрольних рослин; d — шаблон кДНК, що походить від рослин, що зазнали впливу посухи). Ампліфікація геномної ДНК служить додатковим захистом (г). М - маркер довжини ДНК 100 bp (пар основ)
Аналізи зміни експресії генів, що беруть участь в реакції на водний стрес (рослини з тепличного експерименту)
Зміни експресії 13 генів-кандидатів в 11 різних генотипах цукрових буряків спостерігалися на листках рослин, вирощених у тепличних умовах. Зразок експресії, що відповідає BI543243, відрізнявся у рослин, що зазнали впливу посухи, порівняно з відповідними генотипами 1, 10 та 11 в контрольованих умовах (Графік 8). Отже, це може слугувати для розробки молекулярного маркера, корисного для диференціювання генотипів у плані посухи.
Висновки
Стійкість до посухи дуже складний механізм. Експерименти, проведені в трьох різних середовищах (експлантація, тепличні та польові умови) з 11 генотипами, показали, що досить важко визначити єдиний критерій для класифікації генотипів за їх стійкістю до посухи. Однак результати свідчать про те, що накопичення вільного проліну може використовуватися як точний параметр. Класифікація, заснована на зміні концентрації вільного проліну в рослинах, що зазнали впливу посухи в тепличних умовах та під час експлантації, відповідала класифікації, виконаній на основі польових спостережень. Таким чином, подібні дослідження, проведені з молодими рослинами, які, можливо, сприяють застосуванню молекулярних маркерів, можуть бути корисними для визначення селекційного матеріалу щодо стійкості до дефіциту води, що дасть стимул для розвитку селекції цукрових буряків для їх успішного вирощування в умовах майбутніх кліматичних змін.