Позакореневе застосування нанокремнію на цукрових буряках

Цукровий буряк (Beta vulgaris) відомий як рослина, стійка до водного стресу, порівняно з багатьма іншими сільськогосподарськими культурами. Проте на ранніх стадіях розвитку рослини дефіцит води може зашкодити її росту та розвитку. Дефіцит води, що виникає на початкових етапах росту цукрових буряків, може не тільки серйозно перешкоджати розвитку листового апарату, але й меншою мірою обмежувати ріст стрижневого кореня цукрового буряка. Крім того багато досліджень підтверджують, що дефіцит води впливає на деякі фізіологічні та біохімічні ознаки цукрових буряків.
За матеріалами статті на тему «Нанокремній захищає цукрові буряки від водного стресу, покращуючи антиоксидантні системи та сумісні розчини» («Nano‑silicon protects sugar beet plants against water deficit stress by improving the antioxidant systems and compatible solutes») міжнародного журналу Acta Physiologiae Plantarum, вересень 2020 р.
Кремній (Si) вважається одним із найпоширеніших елементів у земній корі після кисню. Хоча кремній дуже рідко зустрічається в ґрунті у вигляді чистих кристалів. Тому польові культури не можуть поглинати достатню кількість кремнію безпосередньо з ґрунту. Корені рослин зазвичай отримують кремній у формі водорозчинної кремнієвої кислоти Si(OH)4, яка зазвичай міститься в ґрунтовому розчині в концентрації 0,1-0,6 ммоль. Дослідники описують позитивний вплив кремнію на ріст і врожайність багатьох польових культур. Однак значущість кремнію для більшості сільськогосподарських культур поки не підтверджена.
Захист, який кремній надає рослинам, може бути пов’язаний з його біохімічною роллю в клітинах та активацією складного антиоксидантного механізму, який покращує стійкість рослин до стресових умов. Кремній може стимулювати накопичення осмолітів, які відіграють життєво важливу роль у підтримці відносного вмісту води листя, що значною мірою сприяє стійкості рослин до посухи.
Крім того, обробка кремнієм проводиться для зменшення токсичного впливу важких металів на польові культури. Також припускається, що кремній може викликати сприятливий вплив на ріст і стійкість рослин до стресу шляхом модифікації їх фізіологічних ознак.
Використання нанотехнологій у сільському господарстві пропонує нові можливості для підвищення врожайності сільськогосподарських культур та зменшення ризиків різних екологічних стресів. Здатність наночастинок захищати рослини від абіотичних стресів залежить не лише від їхньої хімічної структури та реакційної здатності, а й від їх розміру та дози, при якій вони мають максимальну ефективність. Отже, наночастинка може мати різноманітні характеристики.
Незважаючи на те, що були проведені численні дослідження впливу кремнію на пом’якшення стресу сільськогосподарських культур у контрольованих умовах, мало що зрозуміло про роль нанокремнію у зменшенні водного стресу та пов’язаних із цим механізмів у польових умовах. Більше того, більшість досліджень впливу кремнію на польові культури було проведено з використанням силікатних мінералів, таких як силікат натрію, що не може показати реальний вплив чистого кремнію на рослини.
Крім того, до цього часу в статтях не було надано жодного пояснення про роль нанокремнію в накопиченні флавонолів, таких як кверцетин і рутин, у цукрових буряках під час стресу від посухи. Тому дані дослідження мали на меті визначити вплив нанокремнію на ріст цукрових буряків шляхом накопичення осмопротекторів, модифікації газообміну та активації деяких ферментативних і неферментативних антиоксидантів (кверцетину і рутину).
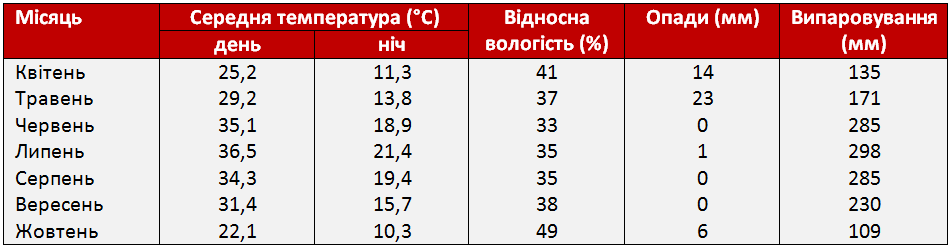
Таблиця 1. Метеорологічні дані вегетаційного періоду
Таблиця 2. Загальна кількість води, використана протягом вегетаційного періоду (I), опади (P), випаровування (ET), обробки
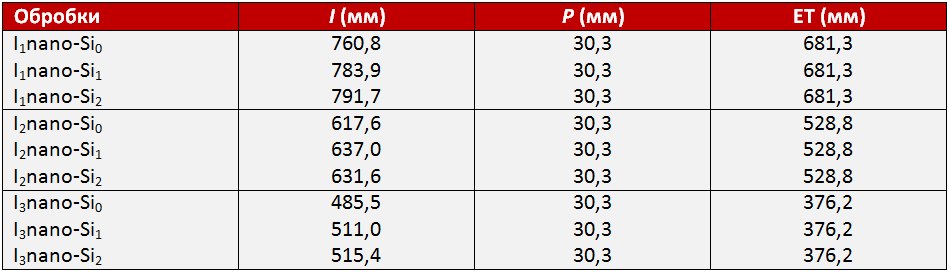
I1 — зрошення при 100% ETC, I2 — зрошення при 75% ETC, I3 — зрошення при 50% ETC, nano-Si0 — 0 ммоль, nano-Si1 — 1 ммоль, nano-Si2 — 2 ммоль.
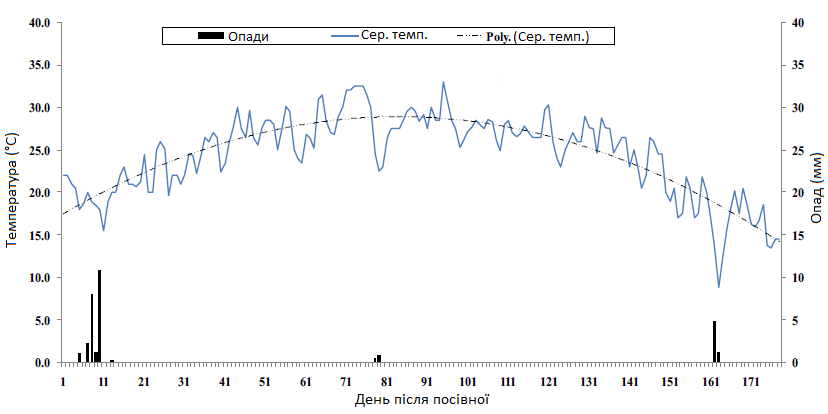
Рис. 1. Середньодобова кількість опадів (мм) та температура повітря (°C) в період вирощування цукрових буряків. Сер. темп.: середньодобова температура в період вегетації; Poly. (Сер. Темп.): поліноміальна лінія тренду, скоригована на середньодобові температури в період росту рослин.
Результати досліджень
Ознаки врожайності та росту
Дефіцит води мав негативний вплив на врожайність і ріст буряків, знизив біомасу, індекс листової поверхні (ІЛП), суху масу пагонів і коренів приблизно на 65%, 31%, 39%, 33% і 21% відповідно. Це показує, що дефіцит води впливає на ріст пагонів більше, ніж на розвиток коренів. Більше того, сильний дефіцит води (50% ETC — евапотранспірація рослин) помітно знизив відносний вміст води в листі (LRWC) на 17%.
Цукрові буряки, оброблені різними дозами нанокремнію, як правило, росли краще, ніж рослини, вирощені без обробки, в умовах повного зрошення або водного стресу. Крім того, застосування нанокремнію в концентрації 1 ммоль на рослинах під дією сильного водного стресу помітно підвищило їх біомасу, урожайність та суху масу пагонів і стрижневого кореня на 21%, 18%, 19% і 7% відповідно, що нижче контрольних рівнів. Однак, після застосовуваня 2 ммоль нанокремнію біомаса, урожайність і суха маса як стрижневого кореня, так і пагонів зменшилися на 41%, 43%, 31% і 48% відповідно. Аналогічно, застосування нанокремнію виявилося однаково корисним для покращення індексу листової поверхні (ІЛП) та відносного вмісту води в листі (LRWC) за всіх режимів зрошення. При слабкому водному стресі (75% ETC) обробка нанокремнієм (I2nano-Si1 та I2nano-Si2) дозволила цукровим бурякам мати ІЛП та LRWC аналогічні до контрольних рівнів. Обробка низькою дозою (1 ммоль) нанокремнію покращила ІЛП та LRWC у рослин під дією сильної посухи, тоді як висока доза (2 ммоль) нанокремнію зменшила ІЛП та LRWC на 54% та 22% по відношенню до контрольних рівнів.
Вміст хлорофілу та параметри, пов’язані з фотосинтезом
Режими зрошення мали менший вплив на вміст хлорофілу в листі, ніж обробки нанокремнієм. Слабкий водний стрес дещо підвищив вміст хлорофілу в цукрових буряках, порівняно з контрольним рівнем. На противагу цьому, при сильному водному стресі спостерігалися помірні тенденції до його зниження. Обробка 1 ммоль нанокремнієм мала адитивний вплив на вміст хлорофілу в рослинах, коли при найвищому рівні водного стресу низька доза нанокремнію (1 ммоль) (I3nano-Si1) показала максимальне значення та збільшила вміст хлорофілу на 12%. Цікаво, що в умовах сильного водного стресу (50% ETC) вміст хлорофілу в рослинах, оброблених 2 моль нанокремнієм, зменшився на 9%, порівняно з контрольним рівнем.
Дефіцит води викликав значне зниження всіх газообмінних параметрів, за винятком чистої швидкості фотосинтезу (PN), яка майже не змінилася за умов слабкого водного стресу. Тим не менш, застосування нанокремнію, як виявилося, пом’якшує інгібуючий ефект, викликаний дефіцитом води. Величина цього пом’якшення була найбільш помітною у рослин, оброблених обома дозами нанокремнію, а також під впливом помірного водного стресу, оскільки показники були подібними до контрольних рівнів (I1nano-Si1). Цукрові буряки, які піддавалися слабкому водному стресу (75% ETC) та оброблені 1 ммоль нанокремнієм, мали чисту швидкість фотосинтезу (PN), продихальну провідність (gs) і транспірацію (T) на 12%, 8% і 3% вищі контрольних рівнів.
Отримані дані показали, що при застосуванні нанокремнію за умови достатнього зрошення всі фотосинтетичні ознаки істотно не змінилися, за винятком чистої швидкості фотосинтезу (PN), яка значно збільшилася під час обробок обома дозами нанокремнію.
Неферментні та ферментативні антиоксиданти
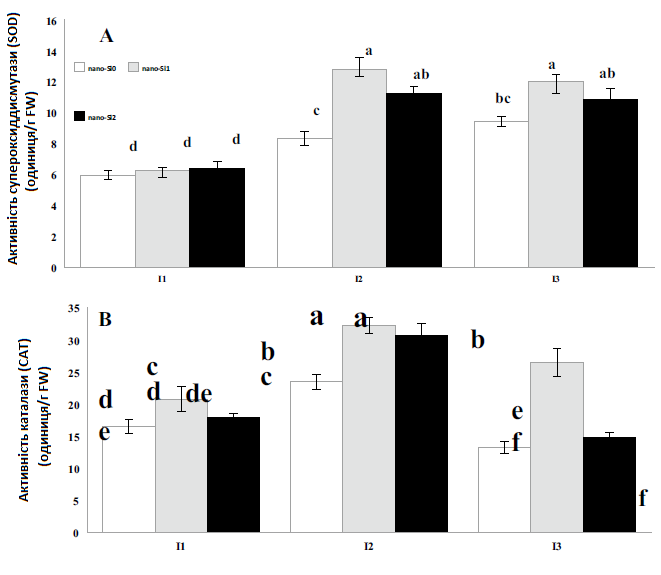
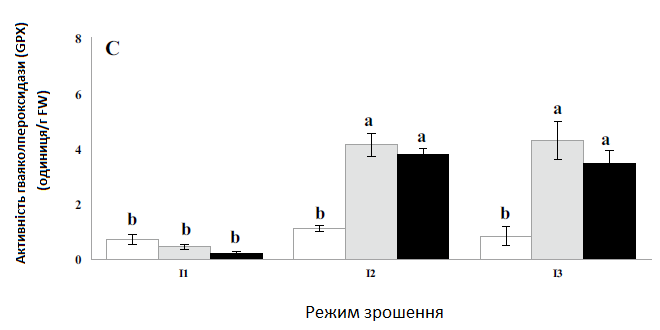
Залежно від дефіциту води можна побачити різні тенденції зміни активності антиоксидантних ферментів, як показано на Рис. 2. Після водного стресу активність супероксиддисмутази (SOD) різко зросла, тоді як активність гваяколпероксидази (GPX) не показала істотних змін у всіх режимах зрошення. У рослин, які відчували помірний водний стрес (75% ETC), активність каталази (CAT) спочатку підвищувалася, а потім при надмірному водному стресі помітно знизилася до нижчого рівня, ніж у контрольних рослин.
Загалом всі вимірювані антиоксидантні ферменти були значною мірою індуковані різними концентраціями нанокремнію (Рис. 2). Найвищі значення активності супероксиддисмутази (SOD) і каталази (CAT) були виявлені при обробці I2nano-Si1, коли рослини перебували під помірним водним стресом (75% ETC) та оброблялися 1 ммоль нанокремнієм. Результати показали, що обробка нанокремнієм збільшила активність супероксиддисмутази (SOD) і каталази (CAT) у 2,1 та 1,9 разів, порівняно з контролем. Вплив водного стресу та нанокремнію був значущим для активності гваяколпероксидази (GPX), проте не було помітної різниці щодо впливу різних доз нанокремнію на активність цього ферменту. Щодо GPX, то при обробці I3nano-Si2 гваяколпероксидаза максимально збільшилася в 6,1 разів, порівняно з контролем.
В умовах сильного дефіциту води (50% ETC) було виявлено значне збільшення загального вмісту фенолу. Сильний дефіцит води підвищив загальний вміст фенолу в 2,4 разів, у порівнянні з контрольними рівнями. Під час слабкого стресу від посухи відмінності в загальному вмісті фенолу не були значущими між не обробленими та обробленими нанокремнієм рослинами. Однак після сильного дефіциту води обробка нанокремнієм (I3nano-Si1 і I3nano-Si2) значно збільшила загальне накопичення фенолу в листі, порівняно з не обробленими рослинами (I3nano-Si0).
Аналіз високоефективної рідинної хроматографії (HPLC) продемонстрував значне збільшення концентрацій рутину та кверцетину під впливом дефіциту води. Сильний водний стрес збільшив кількість кверцетину та рутину в 6 та 2,5 разів, порівняно з контрольними рівнями. Позакореневі обприскування всіма дозами (1 і 2 ммоль) нанокремнію підвищили концентрацію рутину та кверцетину в умовах водного стресу, тим не менш при слабкому водному стресі (75% ETC) не було виявлено статистично значущої різниці між обробленими та необробленими рослинам. Найбільший вміст кверцетину та рутину було виявлено при обробці I3nano-Si2, що продемонструвало збільшення відповідно у 9,5 та 3,9 разів, порівняно з контрольними рослинами.
Рис. 2. Вплив застосування нанокремнію на активність супероксиддисмутази (а), каталази (b) та гваяколпероксидази (c) цукрових буряків за різних режимів зрошення. I1 — зрошення при 100% ETC, I2 — зрошення при 75% ETC, I3 — зрошення при 50% ETC, nano-Si0 — 0 ммоль, nano-Si1 — 1 ммоль, nano-Si2 — 2 ммоль.
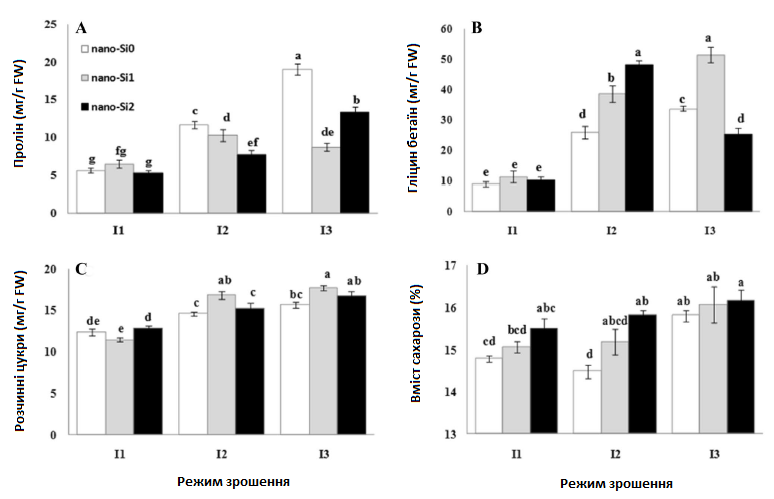
Осмоліти (гліцин бетаїн (GB), пролін і розчинні цукри)
Отримані результати, пов’язані з впливом нанокремнію на осмоліти (гліцин бетаїн (GB), пролін і розчинні цукри) цукрових буряків, зображені на Рис. 3. Висновки продемонстрували загальну тенденцію до більшого накопичення гліцин бетаїну під впливом як нанокремнію, так і дефіциту води. Отримані результати показали, що водний стрес був пов’язаний з кількістю гліцин бетаїну, і якщо посуха продовжувалася, його кількість зростала, максимально збільшившись при I3nano-Si0 в 3,8 разів, порівняно з контрольними рослинами (Рис. 3а). В умовах помірного водного стресу (75% ETC) позакореневе обприскування нанокремнієм (1 і 2 ммоль) було пов’язане зі збільшенням кількості гліцин бетаїну. Однак під час сильного стресу від посухи високий рівень нанокремнію (I3nano-Si2) значно зменшив накопичення гліцин бетаїну, порівняно з низьким рівнем нанокремнію (I3nano-Si1). Максимальне значення гліцин бетаїну було отримано в комбінації 1 ммоль нанокремнію в умовах зрошення I3 (I3nano-Si1) при 51,34 мг/г DW (Рис. 3а).
Вміст проліну в рослинах, які страждали від водного стресу, різко збільшувався. Максимальна кількість проліну, зафіксована при I3nano-Si0, була в 3,4 разів більша, ніж у рослин в умовах оптимального зрошення (Рис. 3b).
Застосування 1 ммоль нанокремнію призвело до різкого зниження вмісту проліну в листі при слабкому (75% ETC) і сильному (50% ETC) стресі від дефіциту води. Однак додавання високої дози (2 моль) нанокремнію значно підвищило вміст проліну, порівняно з відсутньою обробкою в умовах сильного водного стресу (Рис. 3b).
Розчинні цукри показали збільшення приблизно на 18% і 27% при I2nano-Si0 і I3nano-Si0, порівняно з контролем. Застосування нанокремнію підвищило вміст розчинних цукрів в умовах водного стресу. Максимальне збільшення становило близько 43% при I3nano-Si1, порівняно з контрольними рослинами (Рис. 3с).
Помірний водний стрес (75% ETC) призвів до незначного зниження (приблизно на 2%) вмісту сахарози в буряках, тоді як подальше посилення водного стресу до 50% ETC збільшило вміст сахарози приблизно на 7% (Рис. 3d). Застосування нанокремнію не вплинуло на вміст сахарози в рослинах в умовах слабкого водного стресу, тоді як відносне збільшення вмісту сахарози у рослинах, які страждали від сильної посухи, спостерігалося при підвищенні рівня нанокремнію (Рис. 3d).
Рис. 3. Вплив застосування нанокремнію на вміст проліну (а), гліцину бетаїну (b), розчинних цукрів (c) та сахарози (d) у цукрових буряках за різних режимів зрошення. I1 — зрошення при 100% ETC, I2 — зрошення при 75% ETC, I3 — зрошення при 50% ETC, nano-Si0 — 0 ммоль, nano-Si1 — 1 ммоль, nano-Si2 — 2 ммоль.
Окислювальні пошкодження (метилендіоксиамфетамін (MDA) і H2O2)
Швидкість окислювального пошкодження мембрани, оцінена за вмістом метилендіоксиамфетаміну (MDA) та H2O2, була значно вищою в умовах дефіциту води, в порівнянні зі зрошенням. Навпаки, при застосуванні 1 ммоль нанокремнію різке зниження вмісту метилендіоксиамфетаміну (MDA) та H2O2 було виявлено в рослин, які страждали від водного стресу. Цікаво, що в умовах сильного водного стресу (50% ETC) застосування 2 ммоль нанокремнію збільшило вміст метилендіоксиамфетаміну (MDA) без істотних змін у вмісті H2O2. Мінімальна кількість метилендіоксиамфетаміну (MDA) та H2O2 була зафіксована при I1nano-Si2 та I1nano-Si1.
Обговорення
Підтримка активних форм кисню (ROS) в субтоксичній концентрації та покращення водного режиму є основними механізмами захисту рослин, які допомагають впоратися з різними абіотичними стресами, такими як дефіцит води. Останні дослідження показують, що позакореневе застосування нанокремнію може ефективно покращити стійкість рослин до водного стресу, захищаючи фотосинтетичний апарат, покращуючи осмотичне регулювання та зменшуючи окислювальний стрес.
Стрес від дефіциту води посилює вироблення активних форм кисню, порушуючи ланцюги транспорту електронів та окислюючи метаболічні процеси в клітинах рослин. Активні форми кисню (ROS), а саме супероксид-аніон (O2-), гідроксильні радикали (·OH) і перекис водню (H2O2), також можуть негативно впливати на білки, мембранні ліпіди та нуклеїнові кислоти клітин.
Швидкість окислювального пошкодження мембрани оцінюється за вмістом метилендіоксиамфетаміну (MDA) та H2O2. Збільшення виробництва метилендіоксиамфетаміну (MDA) в умовах водного стресу свідчить про серйозні порушення продуктивності та цілісності клітинної мембрани. Тим не менш, додавання низької дози нанокремнію значно зменшило вміст H2O2 та метилендіоксиамфетаміну.
Рослини мають ферментативний антиоксидантний захисний механізм для захисту від активних форм кисню (ROS). Об’єднана робота супероксиддисмутази (SOD), гваяколпероксидази (GPX) і каталази (CAT) необхідна для очищення активних форм кисню (ROS) у рослинах. Металлофермент SOD відомий як перша лінія захисту від пошкоджень, спричинених ROS, і перетворює супероксид-аніон в кисень і перекис водню. Ферменти каталаза (CAT) і гваяколпероксидаза (GPX) діють на H2O2 і розкладають його на кисень і воду.
В умовах стресу від посухи активність активних форм кисню (SOD) підвищується, тоді як активність гваяколпероксидази (GPX) і каталази (CAT) не проявляють аналогічної тенденції. Стрес від дефіциту води суттєво не змінив активність гваяколпероксидази (GPX), проте активність каталази (CAT) посилилася при помірному дефіциті води (75% ETC) і зменшилася при сильному дефіциті води (50% ETC).
Всупереч нашим висновкам, вчені стверджували, що сильний стрес від дефіциту води не має значного впливу на активність каталази (CAT) у листі цукрових буряків. На їх думку, під час водного стресу роль каталази (CAT) була обмеженою. Активність каталази (CAT) чутлива до високого рівня супероксидних радикалів. Таким чином, погіршення її активності може відбуватися через вплив водного стресу на швидкість обороту супероксидних радикалів, що погіршує активність ферменту.
Дане дослідження підтвердило, що листкове обприскування нанокремнієм посилювало ферментативну антиоксидантну активність листя цукрових буряків в умовах посухи. Зниження вмісту метилендіоксиамфетаміну (MDA) та H2O2 можна пояснити Si-залежною ферментативною антиоксидантною активністю.
Попередні дослідження показали, що стрес від дефіциту води може покращити синтез фенольних сполук у відповідь на окислювальний стрес, спричинений накопиченням активних форм кисню (ROS). Фенольні сполуки мають особливу хімічну структуру, яка захищає клітини рослин від окислювального стресу шляхом хелатування металів і зв’язування з вільними радикалами. Поліфенольні молекули, такі як флавоноїди, також пригнічують перекисне окислення ліпідів і покращують стабільність і плинність мембран, діючи як поглиначі вільних радикалів під час стресу від посухи.
У нашому дослідженні дефіцит води викликав різке збільшення вмісту рутину та кверцетину. Проте при слабкому водному стресі загальний вміст фенолу показав незначущу тенденцію до збільшення. Це означає, що рутин і кверцетин можуть бути факторами, викликаними посухою, які відіграють захисну роль в умовах стресу від посухи. Ці результати можна пояснити збільшенням перерозподілу засвоєного вуглецю для біосинтезу фенольних сполук в умовах посухи.
Досі бракує інформації про роль кремнію в накопиченні рутину і кверцетину в польових культурах в умовах стресу від посухи. Попередні дослідження показали зв’язок між вмістом сахарози та фенолу. Таким чином, спричинене кремнієм збільшення розчинних цукрів може сприяти стимуляції виробництва кверцетину та рутину.
У нашому експерименті спостерігалося паралельне збільшення активності каталази (CAT) та вмісту флавонолів (рутину та кверцетину) у листі цукрових буряків під впливом 1 ммоль нанокремнію та дефіциту води.
Здатність продихів закриватися у відповідь на осмотичний стрес є одним із головних механізмів, які дозволяють сільськогосподарським культурам покращувати поглинання CO2 проти втрат води через випаровування. Наші дані чітко показали помітне зниження чистої швидкості фотосинтезу (PN), транспірації (T) та провідності продихів (gs) під впливом водного стресу.
Застосування нанокремнію у низьких дозах значно покращило чисту швидкість фотосинтезу (PN), транспірацію (T) та провідність продихів (gs) як при легкому (75% ETC), так і при сильному (50% ETC) дефіциті води.
Обмежена здатність цукрових буряків регулювати транспірацію та в’янення дорослих і молодих листків свідчать про те, що провідність продихів (gs) і транспірація (T) не є основними обмежуючими факторами поглинання вуглецю за умов недостатнього зрошення.
Основний вплив низького відносного вмісту води в листі (LRWC) на швидкість фотосинтезу викликається неадекватним синтезом рибулози-1,5-бісфосфату (RUBP). Таким чином, більша чиста швидкість фотосинтезу (PN) в рослинах, оброблених нанокремнієм, при сильному дефіциті води (50% ETC) може бути пов'язана з позитивним впливом нанокремнію на гідродинамічну рівновагу в листі цукрових буряків, що підтверджується високою позитивною кореляцією, зафіксованою між відносним вмістом води та швидкістю фотосинтезу. Те, що нанокремній посилює фотосинтез цукрових буряків після водного стресу, можна пояснити збільшенням клітинного виробництва аденозинтрифосфату (ATP).
Незважаючи на те, що більш низький вміст хлорофілу очікувався у рослин, які страждали від водного стресу, як описано щодо різних польових культур, це не спостерігалося у цукрових буряків. Наші дані продемонстрували, що вміст хлорофілу незначно збільшувався при слабкому дефіциті води (75% ETC) і мав тенденцію до зниження при подовженні тривалості стресу від посухи. Невелике збільшення вмісту хлорофілу в умовах слабкого водного стресу може бути пов’язано з меншою питомою площею листя та більшою щільністю клітин на одиницю маси листя цукрових буряків. Крім того, той факт, що параметри газообміну більше реагують на водний стрес, ніж вміст хлорофілу, означає складність взаємозв’язку.
Застосування нанокремнію у концентрації 2 ммоль під впливом сильного водного стресу (50% ETC) значно знизило чисту швидкість фотосинтезу (PN), порівняно з рослинами, які страждали від сильного водного стресу без обробки нанокремніюєм, тоді як вміст хлорофілу та провідність продихів (gs) не змінилися. Ці результати свідчать про те, що обмеження фотосинтетичних процесів, імовірно, пов’язане з не продиховими обмеженнями.
Після обробки нанокремнієм вміст хлорофілу значно збільшується, що можна пояснити впливом нанокремнію на антиоксидантні системи. Була виявлена значна позитивна кореляція між вмістом кверцетину, рутину та хлорофілу в рослинах, оброблених нанокремнієм, що вказує на те, що кверцетин і рутин також можуть діяти як антиоксидант у хлоропластах. Цей результат підтверджує висновки, згідно з яким флавоноїди, розташовані в хлоропластах, допомагають підтримувати активні форми кисню у субтоксичній концентрації.
Під впливом водного стресу осмотична адаптаційна здатність, здається, має особливе значення для тиску тургору та росту рослин. Розчинні цукру та збагачені азотом сполуки, такі як гліцин бетаїн та пролін, є частиною сполук, що накопичуються в цитоплазмі рослинних клітин в умовах посухи, що може бути пов’язано з осмотичною регуляцією. У даному дослідженні водний стрес спричинив значне збільшення осмолітів (цукрів, проліну та гліцин бетаїну) у листі цукрових буряків.
Пролін – це протеїногенна амінокислота, яка накопичується під впливом різних стресів навколишнього середовища і може відігравати вирішальне значення для відновлення рослин після стресу.
У даному дослідженні застосування кремнію значно підвищило вміст гліцин бетаїну, розчинних у листі цукрів та сахарози в цукрових буряках, але зменшило вміст проліну в умовах посухи. Деякі дослідники вважають, що зниження проліну при застосуванні кремнію може бути ознакою зняття стресу.
Останні дослідження показують, що H2O2 бере участь у накопиченні проліну. Ми також спостерігали паралельні зміни рівнів проліну, метилендіоксиамфетаміну (MDA) та H2O2. Ці зміни свідчать про те, що накопичення проліну під впливом водного стресу не обов’язково пов’язане з осмотичним регулюванням і може бути симптомом стресу. Ці висновки дивують, оскільки вони показують, що пролін не може бути індикатором посухостійкості цукрових буряків.
Окрім того, що він є відмінним осмопротектором, пролін виконує кілька функцій: він є джерелом азоту та вуглецю для прискореного відновлення після абіотичних стресів, стабілізатором субклітинних структур, поглиначем вільних радикалів та сигнальною молекулою.
Оскільки вміст гліцин бетаїну, розчинних цукрів та сахарози був значно вищим, ніж вміст проліну в рослинах, оброблених кремнієм, здається, що цукор та гліцин бетаїн можуть відігравати більш помітну роль в осмотичній регуляції. На думку вчених, цукор і пролін, ймовірно, конкурують за вуглецеві скелети.
Одним із цікавих висновків досліджень є те, що під час сильного стресу від посухи (50% ETC) нанокремній на найвищому рівні (2 ммоль) підвищив вміст проліну, що може бути пов’язано з мітохондріальною дисфункцією. Накопичення проліну під час стресу від дефіциту води залежить від поліпшення біосинтезу в хлоропластах і пригнічення деградації в мітохондріях. Добре встановлено, що високі концентрації активних форм кисню, головним чином утворені в мітохондріях, викликають незворотні пошкодження мітохондріальних структур, таких як мембранні ліпіди та білки, що призводить до дисфункції мітохондрій та порушення їхньої роботи. Як показано, високі рівні H2O2 викликають перекисне окислення ліпідів, що підтверджує підвищений окислювальний статус цукрових буряків під час впливу 2 ммоль нанокремнію та сильного стресу від посухи (50% ETC).
Позакореневе застосування нанокремнію стимулювало ріст, збільшувало суху масу, урожайність та біомасу цукрових буряків при всіх режимах зрошення. Кремній може покращувити водний режим рослин, щоб регулювати їх ріст і запобігати старінню листя. Таким чином, причини, що нанокремній збільшує ріст і суху масу в умовах посухи, можуть бути пов’язані з високим вмістом хлорофілу в листі, швидкістю фотосинтезу та покращенням відносного вмісту води в умовах стресу.
Хоча надлишок кремнію зазвичай не вважається шкідливим для рослин, отримані дані показали, що вплив нанокремнію в умовах водного стресу залежав від його дози. Низький рівень (1 ммоль) нанокремнію в умовах сильного водного стресу (50% ETC) був більш сприятливим для сприяння росту та збільшення сухої маси рослин, тоді як високий рівень (2 ммоль) нанокремнію мав шкідливий вплив як на ріст, так і на фізіологічні властивості рослин. Попередні дослідження показали, що наночастинки можуть викликати пошкодження ДНК і загибель клітин. Тому фізико-хімічні властивості наночастинок, такі як розмір і форма, є важливими факторами для визначення потенційної токсичності наночастинок.