Фізіологічна та метаболічна характеристики молодих цукрових буряків в умовах посухи (частина 2)

За матеріалами наукової статті на тему «Фізіологічна та метаболічна характеристики молодих цукрових буряків в умовах тимчасової посухи» («A physiological and metabolic characterization of young sugar beets under temporary drought»), Боннський університет, Німеччина, 2018 р.
Клітинні структури в умовах втрати вологи та відновлення зрошення
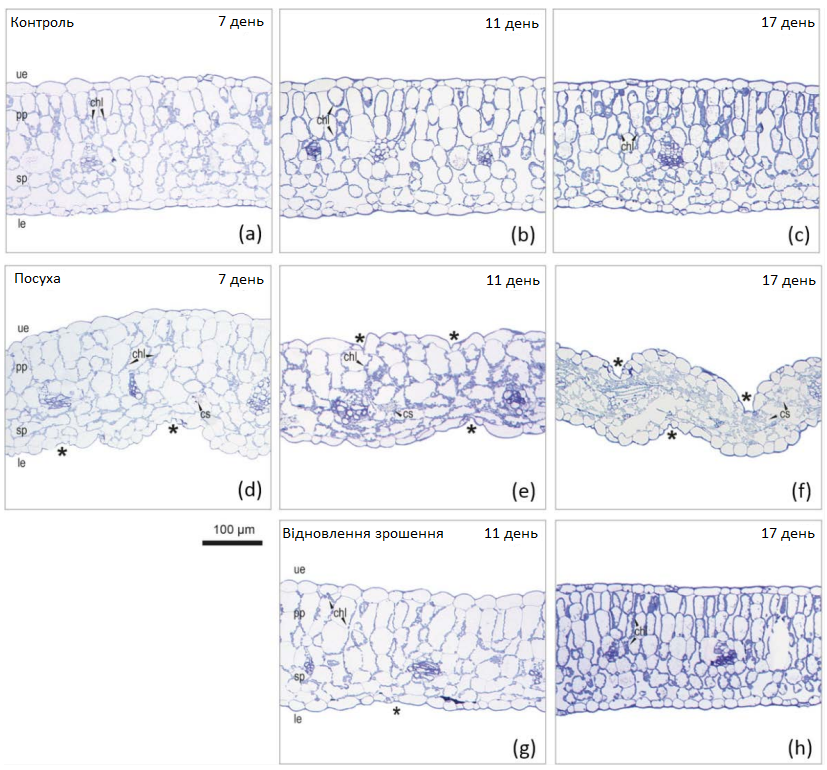
Морфологічні зміни листя цукрових буряків в умовах тривалої посухи та відновлення зрошення були проаналізовані за допомогою мікроскопу (Графік 9). У контрольних умовах палісадна паренхіма листя та клітини зубчастого мезофілу перебували в стані тургору (Графік 9, а). Верхній і нижній епідерміс і восковидна серозна оболонка були плавно зчепленими. Деякі клітини мезофілу демонстрували кристалічні структури (Графік 9, b). В умовах посухи в'янення та втрата тургору призвели до обвисання листя через 7-9 днів (Графік 9, d). Верхній епідерміс та палісадна паренхіма все ще виглядали досить структурованими, але губчастий мезофіл та нижній епідерміс осіли та висохли. Було виявлено зморщені вакуолі, як показано дифузно розташованими хлоропластами. Зі збільшенням дефіциту води в'янення та морфологічні зміни набули більш серйозного характеру, оскільки верхній епідерміс осів та зморщився на 11 день (Графік 9, е). Крім того, кристалічні структури були більш помітними, порівняно з контрольними листками. В останній день посухи весь листок був повністю зім’ятий, а внутрішньоклітинні структури — деформовані (Графік 9, f). Під час відновлення зрошення листя майже повністю відновилося візуально протягом однієї доби (Графік 3, день 11), верхній та нижній епідерміс виглядали гладкими, а палісадна паренхіма та клітини губчастого мезофілу майже відновили свою симетричну структуру (Графік 9, g).Через 9 днів після відновлення зрошення листя повністю відновило запаси вологи, а внутрішньоклітинні структури стали абсолютно симетричними (Графік 9, h).
Графік 9: Найтонші зрізи листя молодих буряків. Найтонші зрізи листя молодих буряків в умовах регулярного зрошення (a-c, контроль), стресу від посухи (d-f) та відновлення зрошення (g, h) на 7, 11 та 17 день експерименту. Зморшкувата тканина вказана як *. Скорочення: cs — кристалічні структури; chl — хлоропласт; pp — палісадна паренхіма; sp — губчаста паренхіма; ue — верхній епідерміс; le — нижній епідерміс.
Зміни метаболітів
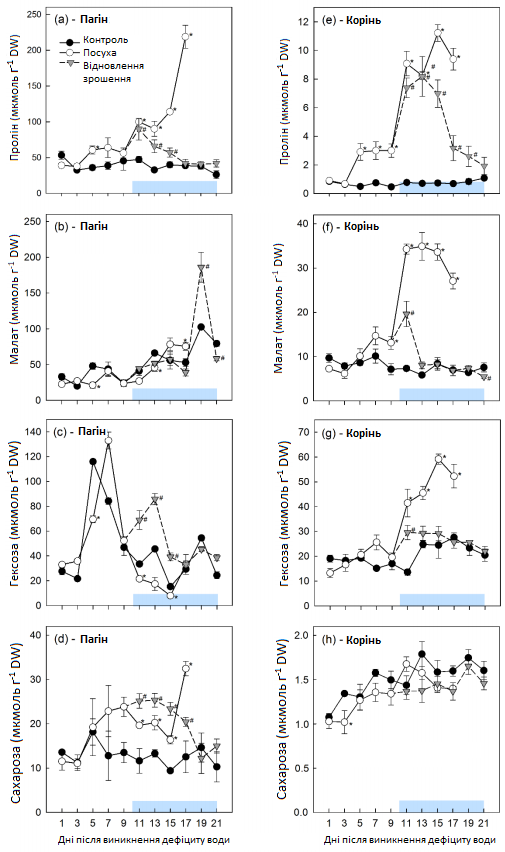
В умовах контролю середні концентрації проліну, малату, гексози та сахарози становили 39,0 ± 7,6, 52,0 ± 24,3, 45,3 ± 30,3 та 12,8 ± 2,3 ммоль г-1 сухої маси (DW) відповідно (Графік 10). У той час як рівень проліну та сахарози залишався постійним, концентрація малату з часом повільно зростала, а концентрації гексози зазнавали коливань і демонстрували сильний пік з 5 по 7 день незалежно від умов вирощування (контроль, посуха, відновлення зрошення). В умовах посухи рівні метаболітів не змінювалися з 1 по 3 день. Після цього рівень проліну повільно збільшувався з 3 по 9 день та інтенсивно — з 9 по 17 день, коли його концентрація досягала максимуму 219 ммоль г-1 DW (Графік 10). Концентрації гексози та малату були нижчими, ніж у контрольних рослинах, з 11 по 13 день із подальшим значним збільшенням рівня малату в 15-17 дні (Графік 10). Концентрація сахарози збільшувалася з 3 по 9 день після припинення зрошення, зменшувалася з 9 по 15 день, а потім знову зросла протягом останніх 2 днів посухи (Графік 10).
Рівні метаболітів у контрольних рослинах відрізнялися між коренями і пагонами, середні концентрації в корені складали 0,7 ± 0,2 ммоль г-1 DW проліну, 7,8 ± 1,3 ммоль г-1 DW малату, 20,3 ± 4,3 ммоль г-1 DW гексози та 1,5 ± 0,2 ммоль г-1 DW сахарози (Графік 10). Через абсорбційну здатність сахарози, її концентрація в корені знаходилася в мілімолярному діапазоні, порівняно з мікромолярними концентраціями в пагоні. Постійне збільшення вмісту сахарози спостерігалося у контрольних рослин, тоді як пролін, малат та гексоза залишалися досить стабільними (Графік 10). В умовах посухи концентрації проліну, малату та гексози не змінювалися з 1 по 3 день, повільно зростали з 3 по 9 день з подальшим різким зростанням (Графік 10). На відміну від листя, вміст сахарози в корені залишався нижчим, ніж у контрольних рослинах, до 9-го дня з подальшим коротким, але значним збільшенням на 11 день, після чого він постійно зменшувався (Графік 10).
Відновлення зрошення призвело до тимчасового збільшення концентрації гексози і сахарози, що перевищило їх концентрації в контрольних рослинах. Концентрації повернулися до контрольних рівнів через 7 (гексоза) та 9 (сахароза) днів відповідно (Графік 10). Збільшення проліну в листі припинилося через 2 дні після відновлення зрошення, його концентрація знизилася до контрольного рівня протягом 7 днів (Графік 10). Концентрації малату повернулися до контрольних рівнів одразу після відновлення зрошення (Графік 10), але максимальна його концентрація спостерігалася 9 днів у період відновлення зрошення. Викликане посухою збільшення рівнів метаболітів у корені припинилося при відновленні зрошення через 2, 4-6 та 5-7 днів відповідно для малату, проліну та гексози (Графік 10). Незважаючи на те, що малат та гексоза повернулися до контрольних рівнів, концентрація проліну залишалася вищою контрольного рівня до кінця всього періоду відновлення зрошення.
Графік 10: Концентрації осмотично активних сполук у пагонах та коренях молодих цукрових буряків. Концентрації осмотично активних сполук у пагонах (a-d) та коренях (e-h) молодих цукрових буряків в умовах регулярного зрошення (контроль, темні кола), стресу від посухи (світлі кола) та відновлення зрошення (трикутники). Горизонтальна пряма являє собою період відновлення. Середні значення ±, n = 4. Кожен день збирання врожаю вказує на значні відмінності для рослин в умовах контролю (α = 0,05): *, P < 0,05 (посуха) та #, P < 0,05 (відновлення зрошення).
Концентрації неорганічних катіонів у коренях
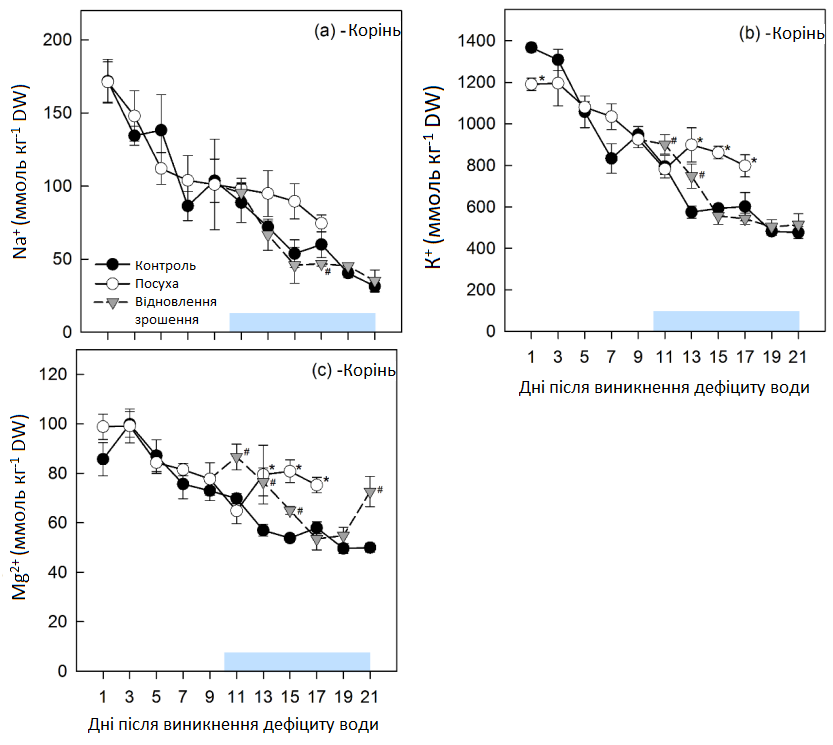
У контрольних рослинах концентрації Na+, K+ і Mg2+ зменшувалися протягом експериментального періоду (Графік 11). В умовах посухи це зменшення сповільнювалося (Na+) або припинялося (K+, Mg2+) протягом останніх 6 днів посухи, а кінцеві концентрації K+ і Mg2+ були значно вищими контрольних рівнів. Після відновлення зрошення концентрації катіонів повернулися до контрольних рівнів протягом 2 (Na+), 6 (K+) та 8 днів (Mg2+) відповідно.
Графік 11: Концентрації неорганічних катіонів у коренях. Концентрації (а) натрію (Na+), (b) калію (K+) та (c) магнію (Mg2+) у коренях молодих цукрових буряків в умовах регулярного зрошення (контроль, темні кола), стресу від посухи (світлі кола) та відновлення зрошення (трикутники). Горизонтальна пряма являє собою період відновлення. Середні значення ±, n = 4. Кожен день збирання врожаю вказує на значні відмінності для рослин в умовах контролю (α = 0,05), позначаються зірочками (посуха) та ромбами (відновлення зрошення).
Внесок метаболітів та неорганічних катіонів в осмотичне регулювання (OA)
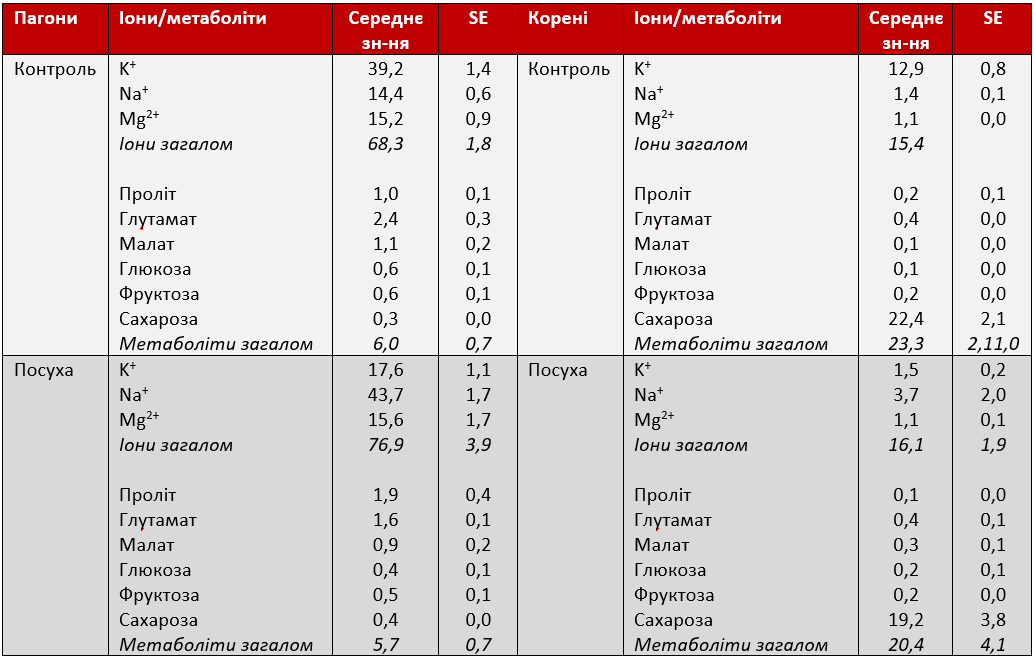
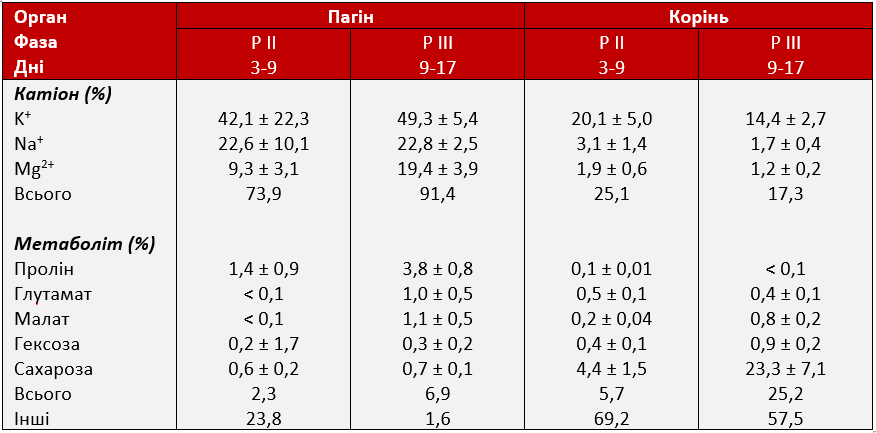
Загальний середній відносний внесок метаболітів та іонів в осмотичний потенціал (OP) між 1 та 17 днями експерименту був різним для пагонів та коренів, але не зазнав істотного впливу стресу від посухи (Таблиця 1). Осмотичний потенціал пагонів залежав переважно від неорганічних іонів (контроль: 68 ± 2%; стрес: 77 ± 4%) та лише незначно від метаболітів (контроль: 6,0 ± 0,7%; стрес: 5,7 ± 0,7%), тоді як у коренях внесок метаболітів був більшим (контроль: 23 ± 2%; стрес: 20 ± 4%), порівняно з іонами (контроль: 15 ± 1%; стрес: 16 ± 2%). В умовах посухи осмотичне регулювання призвело до значного зниження осмотичного потенціалу (OP) з 5-го дня (Графік 6). Для того, щоб визначити, які іони та метаболіти в основному відповідали за осмотичне регулювання, було обчислено відносний внесок кожного іона/метаболіту (∆OP). Осмотичне регулювання пагонів (ОА) між 74% (P II) і 91% (P III) пояснювалося збільшенням концентрації іонів і лише на K+ припадало до 49% OA, потім Na+ (23%) та Mg2+ (19%) (Таблиця 2). Серед метаболітів лише пролін складав 3,8% з 9 по 17 день стресу, тоді як інші метаболіти залишалися нижче 1,5%. У коренях внесок метаболітів в осмотичне регулювання (ОА) з часом збільшувався і сягав 25% протягом 9-17 дня, але внесок кореневих іонів зменшився з 25 до 17% з посиленням стресу (Таблиця 2). Калій знову був основним іоном у корені, але в той час, як пролін був основним метаболітом, що сприяв осмотичному регулюванню (ОА) в пагонах, сахароза відігравала набагато більшу роль у коренях, пояснюючи до 23% ОА. Усі інші метаболіти (включаючи пролін) сприяли ОА менше 1% протягом усього експериментального періоду. Загалом до 98% (у пагонах) та 43% (в коренях) ОА можуть бути пояснені аналізованими іонами та метаболітами.
Таблиця 1: Внесок іонів та метаболітів в осмотичний потенціал листя та коренів цукрових буряків. Показані значення представляють середні дані з 1 по 17 день контрольних рослин та рослин в умовах посухи.
Таблиця 2: Відносний внесок неорганічних іонів та метаболітів в осмотичне регулювання (ОА) пагонів і коренів. Оскільки перше суттєве зниження осмотичного потенціалу (ОP) було виявлено на 5-й день стресу від посухи, внесок обчислювався на 3-9 день (P II: рання захисна метаболічна адаптація) та 9-17 день (P III: посилення метаболічних реакцій та пошкодження клітин), але не на 1-3 день (PI: початкова фаза, що характеризується закриттям продихів листя).
Обговорення
Динамічна послідовність реакцій молодих цукрових буряків на посуху
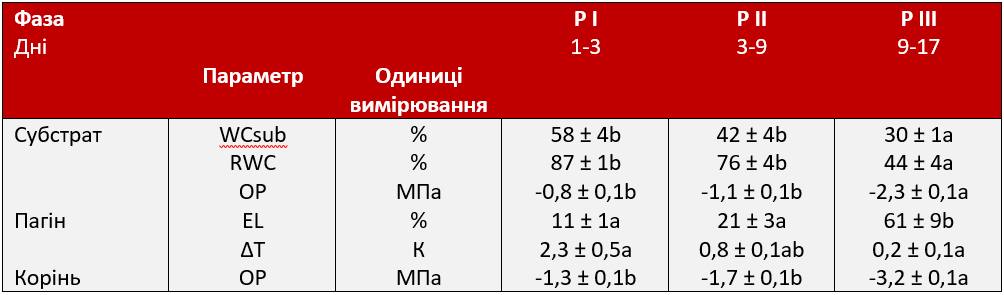
Виділено три фази реакції молодих буряків на посуху на основі водного режиму рослин та ґрунту (RWC, OP, ΔT, WCsub), пошкодження мембрани (EL, Таблиця 3) та метаболітів (проліну, глутамату, малату, гексози, сахарози) (Таблиця 2). Під час початкової фази, починаючи з 1-3 дня (P I), було виявлено лише підвищення температури листя, як первинної фізіологічної адаптаційної реакції. Температура листя є індикатором для транспірації, як було показано, вона дозволяє визначити провідність продихів. Перші ознаки в'янення стали помітними з 3 по 9 день (P II), показники водного режиму рослин та метаболітів значно змінилися, але з повільною швидкістю. Під час третьої фази (9-17 день: P III) фізіологічні зміни посилилися із значно більшою швидкістю та супроводжувалися сильним пошкодженням клітин.
Під час P I та P II не спостерігалося значного пошкодження тканин, хоча спостерігалися втрати води в клітинах зі зменшенням доступності води, що вказувало на початок усадки тканин. P I і P II були чітко розрізнені, оскільки охолоджуючий ефект транспірації листя зменшувався швидко і переважно протягом P I (1-3 день), тоді як метаболічна зміна розпочалася лише в P II (3-9 день). Посуха викликає складну систему біохімічних та молекулярних реакцій, що дозволяють рослинам почати адаптивну реакцію до появи таких візуальних симптомів, як в'янення, до пригнічення росту рослин. Метаболічні зміни в абіотичній реакції на стрес можна розділити на три рівні: (І) миттєва реакція на зміни навколишнього середовища; (ІІ) тимчасова адаптація до стресових умов та (ІІІ) новий режим, встановлений в тривалих стресових умовах.
Таблиця 3. Динаміка стресових реакцій молодих цукрових буряків на посуху. Динаміка стресових реакцій молодих цукрових буряків на посуху, яка характеризується параметрами вмісту води в субстраті (WCsub, виходячи з свіжої маси (FW) субстрату), відносним вмістом води (RWC), осмотичним потенціалом (OP), витоком електроліту (EL) та різницею температур досліджуваного листка та навколишнього середовища (∆T). I фаза (P I) — початкова фаза стресу, що характеризується закриттям продихів; II фаза (P II) — рання захисна метаболічна адаптація; III фаза (P III) — це посилена метаболічна реакція та пошкодження клітин. Щодо кожного параметра, значення, за якими слідує одна і та сама літера, суттєво не відрізняються (α = 0,05).
У цьому дослідженні закриття продихів та припинення транспірації були одними з найбільш ранніх стресових реакцій молодих цукрових буряків на вплив посухи, що передували адаптивним реакціям метаболізму, тому їх можна назвати прямою реакцією на зміну навколишнього середовища. Швидкі зміни рівня ∆T узгоджуються з обмеженою здатністю цукрових буряків регулювати транспірацію в умовах значної потреби в випаровуванні. Швидке закриття продихів вказує на перехід до стратегії економії води рослинами, але також зменшує засвоєння CO2 і може призвести до значного скорочення врожайності. Це особливо згубно у випадках помірних та періодичних умов посухи на початку вегетаційного періоду, коли швидке закриття продихів може представляти небажану ознаку в цільових умовах навколишнього середовища, особливо якщо вони поєднуються з повільним відновленням засвоєння СО2 в умовах відновлення зрошення.
Під час метаболічної адаптації в P II (3-9 днів) рослини, що зазнали впливу посухи, намагалися перейти на новий стійкий рівень, щоб впоратися зі зміною доступності води, що відображається у підвищених рівнях, наприклад, сахарози, малату та проліну. Ця фаза може відповідати тимчасовій адаптації до стресових умов. Значні зміни сахарози, малату або проліну в листі вказують на зміни вуглецевого обміну, який, ймовірно, направляється для підтримки захисних реакцій.
Найбільшою особливістю P III (9-17 дні) було виникнення значного пошкодження мембрани, що свідчило про підвищення рівня витоку електроліту (ELS), що збігалося зі скороченням відносного вмісту води (RWCS) та осмотичного потенціалу пагонів (OPS), першим руйнуванням клітин та деформацією хлоропластів у палісадній та губчастій паренхімі. Відомо, що посуха в кінцевому рахунку призводить до пошкодження клітин через перевиробництво активних форм кисню (ROS), спричинене надмірним скороченням фотосинтетичного ланцюга транспорту електронів та посиленням реакцій Мелера у відповідь на обмеження поглинання CO2 та збільшення фотореспірації. Незважаючи на те, що ні ROS, ні малондіальдегід (продукт деструкції перекисного окислення ліпідів) не були визначені в цьому експерименті, попередні експерименти показали значну кореляцію між витоком електроліту (EL) та концентрацією малодіальдегіду, що вказує на те, що EL можна використовувати як індикатор пошкодження клітин цукрового буряка. Спостережуване значне збільшення пошкодження клітин під час P III вказує на те, що спочатку виробництво ROS врівноважується антиоксидантними реакціями (P II), а потім відбувається пошкодження клітин, коли виробництво ROS перевищує детоксикаційну здатність антиоксидантної системи (P III).
Існує багато свідчень про те, що рослини можуть відчувати ступінь стресу та активувати різні програми реагування на легкі та сильні стреси. Загальна класифікація стресу від посухи на «легкий», «помірний» та «сильний» часто базується на вологоємності ґрунту або потенціалі ґрунтової вологи, але зазвичай бракує інформації про фізіологічний стан рослини, а отже, про очевидний рівень стресу для окремої рослини. Отримані результати свідчать про те, що пошкодження клітин виявляється критичним фактором, що спричиняє перехід цукрових буряків від P II (що представляє "помірний стрес") до P III ("сильний стрес") під час розвитку головного кореня.
Під час останніх 2-х днів стресу від посухи різке збільшення концентрації сахарози та проліну може вказувати на інше порушення метаболізму, можливо, пов’язане з початком процесу старіння, спричиненого накопиченням активних форм кисню (ROS) в умовах сильного стресу від посухи. Загально відомим є співвідношення між накопиченням цукру та старінням листя. Незважаючи на накопичення сахарози в пагонах під час пізньої фази стресу, внесок в осмотичну регуляцію (ОА) був незначним (< 1%, Таблиця 1), що підтверджує думку про те, що сахароза може функціонувати як сигнальна молекула або стабілізатор мембрани, а не як осмопротектор у пагонах цукрових буряків. В умовах дефіциту води спостерігалося значне накопичення проліну в цукрових буряках, що може розглядатися як ще один показник старіння, що виникає внаслідок розпаду білка, а також як захисний механізм, що сприяє стабілізації мембрани.
Осмотичне регулювання в коренях цукрових буряків
Коренеплоди — органи накопичення поживних речовин, що характеризуються високим накопиченням сахарози до 19% від свіжої маси (FW) кореня під час збирання. Даний експеримент було проведено, коли коренеплоди тільки но починали накопичувати сахарозу, на що вказувало повільне, але постійне збільшення її вмісту в контрольних коренеплодах. Висока концентрація сахарози дозволяє цукровим бурякам встановити набагато нижчий осмотичний потенціал (OP) у коренях, порівняно з пагонами, що частково може відповідати за здатність компенсувати тимчасовий дефіцит води. Разом із K+, сахароза була одним із головних «учасників» осмотичної регуляції (ОА) в коренях. Однак під час посухи концентрація сахарози в корені не зростала в достатній мірі, що повністю пояснює спричинене посухою значне скорочення осмотичного потенціалу кореня (OPR). Інші сумісні осмоліти також збільшувалися під час P III, але їх концентрація була низькою, порівняно з пагонами, а їх здатність сприяти осмотичному регулюванню (ОА) була обмеженою, на що також вказують стабільні показники проліну та малату з 11 по 15 день, коли OPR все ще продовжував знижуватися. Незважаючи на те, що ці метаболіти не суттєво сприяли осмотичній регуляції (ОА) в коренях, їх збільшення, викликане дією стресу, все ще свідчить про інші важливі функції, як от наприклад захист від активних форм кисню та, отже, від пошкодження мембрани. Оскільки внесок катіонів в осмотичну регуляцію в коренях також зменшився з часом, потрібні додаткові дослідження для виявлення інших сполук, які підтримують 58-69% осмотичної регуляції (ОА) в коренеплодах в умовах стресу. Метаболіти, які, як повідомлялося раніше, збільшуються в коренеплодах в умовах посухи, — це гліцинбетаїн та глютамін, не були враховані в даному дослідженні, але можуть відігравати важливу роль в осмотичному регулюванні (ОА) в коренях цукрових буряків.
Динаміка процесу відновлення після тимчасового стресу від посухи
Незважаючи на те, що цукрові буряки, вирощувані в польових умовах, мають гарну здатність до відновлення після тимчасових посушливих періодів, майже нічого не відомо про основні реакції метаболізму та наскільки вони відновлюються після посухи. У даному дослідженні відновлення зрошення починалося, коли в листі почав посилюватися мембранний потік, а пошкодження клітин було підтверджене гістологічним аналізом. В’янення та пересихання тканин молодого листя зникло протягом одного дня після відновлення зрошення, але водний режим та зміни метаболізму для осморегуляції потребували більше часу для повернення до контрольних рівнів. Ріст пагону та кореня відновився лише після того, як відносний вміст води (RWC) та осмотичний потенціал (OP) обох органів майже досягли контрольних рівнів, що свідчило про відновлення достатнього тургору, необхідного для відновлення клітин.
Протягом перших 6 днів після відновлення зрошення концентрації гексози та сахарози в листі перевищили контрольні рівні, що свідчить про те, що фотосинтетична активність була негайно, принаймні частково, відновлена. Однак ріст все ще гальмувався, тому накопичувальна здатність пагонів та коренів була низькою, що, ймовірно, призводило до накопичення розчинних цукрів у тканинах. Цукри повернулися до контрольного рівня приблизно в той же час, коли відновився ріст, що вказує на те, що вони слугували джерелом енергії та полегшили початок росту рослин після тимчасового посушливого періоду. Концентрації проліну, які постійно зменшувалися, вказують на його втрату та, можливо, його використання в якості альтернативного джерела енергії під час початкової фази відновлення.
Щонайменше 9 днів потрібно ∆T для повернення до контрольного рівня та адаптації до зростаючої доступності води, навіть, незважаючи на те, що сітчасті структури виявилися повністю функціональними, а відносний вміст води в пагонах (RWCS) повернувся до контрольного рівня через 7-8 днів. Це відповідає даним про постійне зменшення фотосинтетичного поглинання СО2 після тимчасового водного стресу. Фізіологічні обмеження відновлення фотосинтезу після посухи до кінця не вивчені, але повільне відновлення клітинних мембран, як вказує ELS, підтримує думку про те, що фотосинтетичне перенесення електронів у хлоропластах може залишатися порушеним протягом більш тривалого періоду через пошкодження мембрани. Постійне (часткове) закриття продихів, яке спостерігалося у різних видів рослин і тривало до декількох тижнів після відновлення зрошення, може бути альтернативним поясненням спостережуваного повільного відновлення процесу транспірації. Була висунута гіпотеза, що стійке закриття продихів листя може бути фізіологічним регулюванням для запобігання подальших втрат води в очікуванні майбутніх стресових умов. Скорочення поглинання СО2 та додаткове обмеження росту через стійке закриття продихів можна зменшити шляхом використання активності малік-ферментів, забезпечуючи збільшення концентрації СО2 в листі. У даному дослідженні швидке зменшення концентрації малату в коренях в умовах відновлення зрошення та максимальний перехідний рівень малату в пагонах можна трактувати як перенесення малату від кореня до листя, де він може служити додатковим джерелом CO2.
Фаза затримки відновлення осмотичного потенціалу (OPR), концентрацій проліну та малату в коренях вказує на те, що налагодження адаптивних метаболічних реакцій у коренях відбувається повільніше, ніж у пагонах. Це має фізіологічний сенс, оскільки в польових умовах середовище, в якому функціонує коренева система рослини, менше піддається впливу швидко мінливих умов навколишнього середовища, як наприклад посуха зазвичай розвивається повільніше в ґрунті, порівняно з повітрям. Повільне припинення метаболічних реакцій дозволило б «підготувати» коренеплід до наступного посушливого періоду, що може бути механізмом енергозбереження за несприятливих умов.
Вихід цукру залежить від заміщення нецукрових сполук сахарозою під час росту головного кореня при збалансованому постачанні води та живильних речовин, і було висунуто припущення, що накопичення сахарози, ймовірно, обмежується підвищеним рівнем осмолітів або катіонів, таких як K+, Na+ та Mg2+. Це узгоджується з наведеними даними, коли максимальні рівні сумісних осмолітів та підвищені концентрації катіонів під час P III співпадали з постійним зниженням концентрації сахарози в коренеплоді. Крім зниження виходу цукру, високі рівні осмолітів та іонів додатково погіршують технічну якість коренеплодів, обмежуючи кристалізацію цукру. У польовому дослідженні після 35 днів стресу від посухи 2 місяці процесу відновлення виявилося недостатньо для запобігання накопичення Na+ та α-аміно-азотних сполук. Однак у даному дослідженні концентрації осмолітів та катіонів у коренеплодах повернулися до контрольних рівнів протягом 8 днів після відновлення зрошення, що вказує на те, що тимчасові стреси від посухи під час раннього розвитку цукрових буряків навряд чи матимуть негативний вплив на технічну якість сировини, якщо тільки посуха не проявиться на заключному етапі розвитку цукрового буряка.
У цілому, наведені результати свідчать про те, що проаналізований сорт Паулета майже повністю відновився протягом періоду відновлення зрошення (11 днів) після 9 днів посушливого періоду, який викликав зміни водного режиму, а також значне пошкодження мембрани. Однак різні параметри вимагають різних проміжків часу для повного відновлення та повернення до контрольних рівнів, що може бути доречно, якщо рослини стикаються з неодноразовими посушливими періодами лише з короткими проміжками часу для відновлення. Дане дослідження також вказує на те, що одна тимчасова посуха на ранній стадії розвитку молодих цукрових буряків може вплинути на подальший розвиток рослин не лише тимчасовим пригніченням росту коренеплодів та пагонів під час стресового періоду, а й зменшенням поглинання CO2, яке триватиме, навіть, після завершення посухи та відновлення зрошення. Чи вплине це на кінцеву врожайність, або можливо буде компенсовано протягом решти періоду росту, ймовірно, залежатиме від подальшої доступності води.
Перевага поєднання інфрачервоної термографії (ІRТ) та деструктивних аналізів
Вчасне виявлення ранніх стадій стресу є надзвичайно важливим у селекційних програмах, де велика кількість культурних сортів рослин потребують характеристики в умовах високої пропускної здатності. IRT дозволяє здійснювати неінвазивний високопродуктивний контроль рослин в умовах обмеженої доступності води в польових та тепличних умовах. Однак придатність інфрачервоної термографії виявляти дуже ранні реакції на посуху ще не була продемонстрована. Аналогічно, інвазивні вимірювання не користуються успіхом у поєднанні з недеструктивними підходами для виявлення початкових реакцій на стрес, і навіть незважаючи на те, що вони можуть бути адаптовані до умов високої пропускної здатності за допомогою роботизованих платформ, вони все ще вимагають багато часу і не дозволяють проводити безперервні вимірювання. У даному дослідженні було показано, що IRT дійсно дає змогу відслідковувати початкове зменшення транспірації після виникнення стресу від посухи до виявлення значних змін за допомогою деструктивних вимірювань. Отже, IRT може бути перспективним інструментом для швидкого виявлення початкових реакцій на стрес великої кількості генотипів і може прискорити широкомасштабне високопропускне фенотипування в контрольованих умовах навколишнього середовища. Однак окремо IRT не в змозі розрізнити пізні фази стресу, тому не може виокремлювати генотипи, що відрізняються своєю реакцією на сильніший стрес від посухи. Слід бути обережними при перенесенні результатів із тепличних досліджень на польові умови, оскільки на температуру рослин у полі впливають різні зовнішні фактори, такі як архітектура рослин, температура ґрунту та погодні умови. Однак використання технології IRT в польових умовах постійно вдосконалюється і може стати цінним інструментом для попереднього симптоматичного виявлення стресу від посухи та оптимізованої організації зрошення, наприклад, у високопродуктивних зрошувальних сільськогосподарських системах.
Недеструктивні методи можуть оцінити генотипічні відмінності реакцій рослин, наприклад раннє закриття продихів листя після виявлення стресу або час, необхідний для відновлення, але вони дають обмежену інформацію про основні метаболічні процеси чи генетичні ознаки. Неінвазивні вимірювання не можуть визначити те, чи є повільне відновлення транспірації генотипною ознакою, яка забезпечує своєрідний "прогностичний захист" від майбутніх посушливих періодів, або воно є лише відповіддю на ступінь пошкодження клітин фотосистеми. Поєднуючи неінвазивні та деструктивні вимірювання, дане дослідження, схоже, вказує на те, що повільне відновлення транспірації корелює з пошкодженням клітин (ELS). Це може виявити можливість використовувати неінвазивні вимірювання відновлення ІRТ для відбору перспективних генотипів, на які менше впливає пошкодження клітин під час попереднього стресу від посухи.
|
Висновки Інфрачервона термографія (ІRТ) була придатною для виявлення початкового зменшення транспірації, спричиненого посухою, до того, як значні зміни були визначені за допомогою інвазивного (метаболічного або фізіологічного) аналізу. Однак лише одна IRT не може розрізняти більш важкі фази стресу. Під час безперервного стресу від посухи у молодих цукрових буряків спочатку спостерігається закриття продихів листя, яке супроводжується фазою захисної метаболічної адаптації та, нарешті, посилюються метаболічні реакції, що збігаються зі збільшенням ушкоджень клітин. Хоча більшість метаболічних параметрів повернулися до контрольних рівнів протягом 10 днів після відновлення зрошення, транспірація листя та пошкодження мембрани відновлювалися досить повільно. Це може мати тривалий вплив на поглинання СО2 протягом решти періоду росту і, таким чином, на кінцеву врожайність та/або підтримку готовності рослин краще справлятися з наступними посушливими періодами. Для селекційних цілей швидкість і завершеність процесу відновлення може стати корисною ознакою. Поєднання неінвазивних (ІRТ) методів фенотипування з аналізом відібраних метаболітів може прискорити відбір сортів, краще адаптованих до посушливих умов. |