Фізіологічна та біохімічна реакція цукрових буряків на вплив ультрафіолетового випромінювання (частина 1)

Ультрафіолетове випромінювання може спричиняти багато серйозних проблем для всіх живих організмів, а чутливість сільськогосподарських культур до УФ-опромінення вимагає особливої уваги. Для визначення придатності цукрових буряків (Beta vulgaris L) до вирощування в районах з УФ-опроміненням було досліджено вплив різних доз УФ-випромінювання (3,042, 6,084 та 9,126 кДж-2d-1). Ультрафіолетове випромінювання викликає значне сповільнення розвитку рослин, що відображається як у зменшенні їх росту, так і свіжої та сухої маси. Однак, це не залежало від УФ-дози, а було пов'язано зі зменшенням утворення фотосинтетичного O2, відносного вмісту хлорофілів, фотосинтетичних пігментів та флуоресценції хлорофілу. З іншого боку, активність антиоксидантних ферментів, загальний вміст білка, сумісні осмоліти, загальний вміст вільних амінокислот та загальний вміст беталаїну збільшилися під дією 9,126 кДж-2d-1 ультрафіолетового випромінювання, що представляє механізми, завдяки яким рослині вдалося впоратися зі стресом. Оксидативний стрес проявився у збільшенні вмісту малондіальдегіду (MDA), проте перекис водню (H2O2) не зазнав жодного впливу. Таким чином, досліджуваний сорт цукрових буряків BR1 вважається придатним для вирощування в районах із високими дозами ультрафіолетового випромінювання.
За матеріалами статті на тему «Фізіологічна та біохімічна реакції цукрових буряків (Beta vulgaris L) на ультрафіолетове випромінювання» («Physiological and biochemical responses of sugar beet ( Beta vulgaris L) to ultraviolet-B radiation») Паріса Рахімзаде Карвансара та Сеїда Мехді Разаві, Факультету біології, Кафедри природничих наук, Університету Мохаггег Ардабілі, м. Ардебіль, Іран.
Ультрафіолетове випромінювання (УФ) з довжиною хвилі від 10 нм до 400 нм становить близько 10% від загальної віддачі сонячного світла. Випромінювання високої енергії в основному поглинається киснем та озоном в атмосфері. Тому, руйнування озонового шару, яке випадково було виявлено в 90-х роках ХХ століття, пов'язане переважно з підвищенням рівня ультрафіолетового випромінювання та, відповідно, його впливом на здоров'я людини. У різних куточках нашої планети поточні рівні УФ-випромінювання коливаються в межах від 2 до 12 кДж-2 на добу. Ультрафіолетове випромінювання також може порушувати та гальмувати ріст і розвиток рослин, ускладнювати процес фотосинтезу, розклад фітомаси, темнове дихання та транспірацію. Це є наслідком пошкодження макромолекул УФ-випромінюванням, що спричиняє розриви подвійної спіралі ДНК, перекисне окислення ліпідів та руйнування білка у рослині. Однак рослини можуть проявляти захисні реакції та досягти певного ступеня стійкості до УФ-випромінювання.
Стійкість цукрових буряків до ультрафіолетового випромінювання змінюється відповідно до різних сортів та, навіть, тісно пов'язаних генотипів. Однією з найпоширеніших реакцій рослини на вплив ультрафіолетового випромінювання є зменшення листової поверхні як через обмеження цитокінезу, так і через ріст клітин. З іншого боку, рослини проявляють біохімічну та фітохімічну адаптацію до впливу ультрафіолетового випромінювання шляхом посилення їх первинних та вторинних метаболітів для підвищення ефективності захисту. Одним класом вторинних метаболітів, які захищають від УФ, є фенольні сполуки, такі як лігнін — компонент клітинної стінки всіх судинних рослин та деяких водоростей. Здерев’яніння клітинної стінки відбувається під впливом УФ-випромінюванням і є першим захисним бар'єром епідермісу від УФ-променів. Серед інших фенілпропаноїдів, ефективних у захисті від ультрафіолетового випромінювання, найбільш поширеними є антоціани. Цікаво, що більшість рослин порядку гвоздикоцвіті (лат. Caryophyllales) містять беталаїни замість антоціанів. Беталаїни — це клас сполук з азотистим фібрилярним центром, які виконують функцію антиоксидантів у рослинах та в харчуванні людини. Ці дві групи пігментів ніколи не зустрічаються в одних і тих же рослинах, оскільки ферменти, що продукують антоціани, не виявляються в рослинах, які містять беталаїни.
Однією з рослин, які виробляють беталаїни, є цукровий буряк (Beta vulgaris), який завдяки своїй здатності накопичувати велику кількість цукру в корені забезпечує близько 40% світового виробництва цукру. Однак негативний вплив навколишнього середовища може зменшити потенціал виробництва цукру шляхом погіршення продуктивності цукрових буряків. Було проведено багато досліджень з метою визначення реакції цукрових буряків на абіотичні стреси, включаючи посуху, спеку, холод та УФ-випромінювання. Дійсно, під впливом УФ-випромінювання на фоні різних умов освітлення було виявлено зменшення листової поверхні, свіжої та сухої маси, вмісту хлорофілів та каротиноїдів у цукрових буряках. Крім того, раніше повідомлялося, що ультрафіолетове випромінювання знижує флуоресценцію хлорофілу у 22 комерційних сортах цукрових буряків. Ці результати показали, що швидкість відновлення була різною у всіх сортів, особливо між найбільш чутливими та найбільш стійкими сортами буряків. Як правило, попередні дослідження довели, що цукровий буряк є стресостійкою культурою.
У даній статті було досліджено широко використовуваний іранський багаторічний сорт цукрових буряків BR1 та його фізіологічну та біохімічну реакції на три різні дози УФ-випромінювання, щоб оцінити його потенціал як ефективної культури в районах з високими дозами УФ-випромінювання.
Матеріали та методи
Умови вирощування та вплив УФ-випромінювання
 Насіння сорту BR1 цукрового буряка (Beta vulgaris L.) (отримане в Організації сільського господарства провінції Західний Азербайджан, Урмія, Іран) оброблялося 10% гіпохлоритом натрію протягом 10 хв і потім замочувалося у дистильованій воді. Після 4 днів проростання розсаду пересаджували в горщики. Всі рослини вирощувалися у кліматичній камері (Jeiotech GC-3007 LH, Корея) при 25/20 °C (цілодобово), при 16-годинному світлому / 8-годинному темному фотоперіоді протягом 30 днів при інтенсивності світла 500 мкмоль m-2s-1. Після цього рослини були розділені на 4 групи, включаючи контрольну та три експериментальні групи, що піддавалися ультрафіолетовому випромінюванню 3,042, 6,084 та 9,126 кДж-2d-1 протягом одного тижня. УФ-випромінювання забезпечувала лампа TL 20W/12 RS SLV/25 (Philips, Німеччина), розташована над рослинами на відстані 20 см. Лампа була загорнута в 0,13 мм діацетилцелюлозну та поліетиленову плівку для поглинання випромінювання нижче 290 нм (УФ-С) та вище 320 нм (УФ-А), відповідно. Інтенсивність випромінювання вимірювалася за допомогою спектрорадіометра ASD Handheld 2.
Насіння сорту BR1 цукрового буряка (Beta vulgaris L.) (отримане в Організації сільського господарства провінції Західний Азербайджан, Урмія, Іран) оброблялося 10% гіпохлоритом натрію протягом 10 хв і потім замочувалося у дистильованій воді. Після 4 днів проростання розсаду пересаджували в горщики. Всі рослини вирощувалися у кліматичній камері (Jeiotech GC-3007 LH, Корея) при 25/20 °C (цілодобово), при 16-годинному світлому / 8-годинному темному фотоперіоді протягом 30 днів при інтенсивності світла 500 мкмоль m-2s-1. Після цього рослини були розділені на 4 групи, включаючи контрольну та три експериментальні групи, що піддавалися ультрафіолетовому випромінюванню 3,042, 6,084 та 9,126 кДж-2d-1 протягом одного тижня. УФ-випромінювання забезпечувала лампа TL 20W/12 RS SLV/25 (Philips, Німеччина), розташована над рослинами на відстані 20 см. Лампа була загорнута в 0,13 мм діацетилцелюлозну та поліетиленову плівку для поглинання випромінювання нижче 290 нм (УФ-С) та вище 320 нм (УФ-А), відповідно. Інтенсивність випромінювання вимірювалася за допомогою спектрорадіометра ASD Handheld 2.
Визначення відносного вмісту води в листі та висоти рослини
Висота рослини, свіжа маса та листова поверхня визначалися відразу після збирання буряків. Суха маса визначалася після висихання при 75 °C протягом 48 год. Відносний вміст води визначався у наймолодшому листочку головного пагона. Після визначення свіжої маси (FW), листя поміщали в темну кімнату в дистильовану воду на 24 години з метою визначення маси при насиченні водою (TW). Потім зразки висушувалися при 75 °C для визначення сухої маси (DW). Відносний вміст води у листі розраховувався за наступною формулою:
LRWC(%) = [(FW − DW)/(TW − DW)] × 100.
Визначення фотосинтетичних пігментів
Для визначення вмісту хлорофілів та каротиноїдів 0,1 г зрілих листків подрібнювали в 1 мл 80% ацетону та центрифугували при 2700 об./хв протягом 10 хв. Надосадова рідина безпосередньо використовувалася для спектрофотометричних вимірювань поглинання при 647, 663 та 470 нм. Для визначення хлорофілу a, хлорофілу b, загального вмісту хлорофілів та каротиноїдів за допомогою спектрофотометра використовувались такі рівняння:
Загальний вміст хлорофілів (мкг/мл) = (20,21 × A645) + (8,02 × A663)
Хлорофіл a (мкг/мл) = (12,25 × A663 – 2,79 × A647)
Хлорофіл b (мкг/мл) = (21,50 × A647 – 5,10 × A663)
Каротиноїди (мкг/мл) = (1000 × A470 – 1,82chlorophyll a – 85,02chlorophyll b)/198.
Визначення зміни фотосинтетичного O2
Для визначення зміни фотосинтетичного O2 було використано диференціальний аналізатор O2 S104 Qubit Systems (DOX). Кліматичну камеру відкалібрували за допомогою азотного газу, а потім додавали до 21% кисню (O2). Зразки листя поміщали в камеру, яку наповнювали атмосферним повітрям. Виділений з листя кисень вимірювали сенсорним пристроєм протягом 1000 с. Світло було на рівні 66 мкмоль m-2s-1.
 Максимальний квантовий вихід ФСII та визначення відносного вмісту хлорофілів
Максимальний квантовий вихід ФСII та визначення відносного вмісту хлорофілів
Перед збиранням буряків визначалася концентрація хлорофілів у кожному четвертому листу усіх рослин за допомогою вимірювача вмісту хлорофілів (Hansatech Instruments, CL-01, Великобританія).
 Максимальний квантовий вихід ФСII (Fv/Fm) (Fv — змінна флуоресценція; Fm — максимальна флуоресценція) визначався перед збиранням за допомогою аналізатора продуктивності рослин (Hansatech Instruments, Handy PEA, Великобританія).
Максимальний квантовий вихід ФСII (Fv/Fm) (Fv — змінна флуоресценція; Fm — максимальна флуоресценція) визначався перед збиранням за допомогою аналізатора продуктивності рослин (Hansatech Instruments, Handy PEA, Великобританія).
Визначення вмісту вільних амінокислот, проліну та гліцин бетаїну
Для визначення концентрації вільних амінокислот свіжий зразок гомогенізували в 50 ммоль фосфатному буфері (pH 6,8). Екстракти центрифугували протягом 20 хв при 3000 об./хв, потім до супернатанту додавали нінгідриновий реагент і нагрівали протягом 7 хв на водяній бані при 100 °С. Коефіцієнт поглинання при 570 нм визначався відразу після охолодження на холодній водяній бані. Для побудови калібрувальної кривої використовували різні концентрації гліцину. Для оцінки вмісту проліну 0,5 г тканин свіжого листя подрібнювалися з додаванням 10 мл сульфосаліцилової кислоти. Після додавання 2 мл нінгідринового реагенту, 2 мл оцтової кислоти та 2 мл толуолу з'явились дві різні фази. Коефіцієнт поглинання забарвленого супернатанту, що містив толуол та пролін, вимірювали при 520 нм. Калібрувальна крива була побудована з використанням різних концентрацій проліну. Для визначення концентрації гліцин бетаїну 0,5 г тканин висушеного листя подрібнювалися, гомогенізувалися з 20 мл дистильованої води і струшувалися протягом 24 годин при 25 °С. Після фільтрування зразки розбавлялися у пропорції 1:1 з 2N сірчаною кислотою, охолоджувалися протягом 1 години і додавалося 10 мл розчину Люголя. Після інтенсивного перемішування всі зразки центрифугували і зберігали при 4 °С протягом 16 годин. Залишкові кристали розчиняли в 1, 2-дихлоретані, а коефіцієнт поглинання визначався при 365 нм. Калібрувальна крива будувалася з урахуванням різних концентрацій гліцин бетаїну в 1N H2SO4.
Визначення рівня перекису водню (H2O2)
Для визначення вмісту H2O2 тканини свіжого листя подрібнювалися з 1% трихлороцтової кислоти. Зразки центрифугували при 4 °С протягом 15 хв при 10,000 г. 0,5 мл супернатанту змішували з 0,5 мл 10 ммоль фосфатного буфера (pH 7) і 1 мл 1 моль йодиду калію. Вміст H2O2 визначався спектрофотометрично при 390 нм. Для калібрувальної кривої використовували різні концентрації H2O2.
Екстракція білка
0,05 г тканини свіжого листя гомогенізували з 2 мл 0,1 моль фосфатного буфера (pH 6,8). Всі екстракти центрифугували при 13,000 г протягом 15 хв при 4 °С і використовували для визначення активності ферментів та вмісту білка. Концентрація білка визначалася за допомогою методу Бредфорда з використанням альбуміну бичачої сироватки.
Визначення активності каталази (CAT)
Для дослідження активності каталази було підготовлено реакційну суміш, яка включала 2,5 мл 0,05 моль фосфатного буфера (pH 7), 0,3 мл 3% H2O2 та 0,2 мл екстракту білка. Ферментативна активність досліджувалася за методом Бера — Ламберта як видалення H2O2, виміряне за рахунок зменшення коефіцієнта абсорбції при 240 нм.
Визначення активності поліфенолоксидази (РРО)
Для визначення активності поліфенолоксидази реакційну суміш, що включала 2,5 мл 0,2 моль фосфатного буфера (pH 6,8) та 0,2 мл 0,02 моль пірогалолу, витримували на водяній бані при 40 °С. Після додавання 0,2 мл ферментного екстракту визначалися зміни коефіцієнта абсорбції при 430 нм. Ферментативна активність визначалася за допомогою методу Бера — Ламберта.
Визначення активності аскорбатпероксидази (APX)
Для визначення активності аскорбатпероксидази реакційна суміш включала 2,5 мл 0,2 моль фосфатного буфера (pH 6,5), 0,2 мл 3% H2O2, 5 ммоль аскорбінової кислоти та 0,2 мл ферментного екстракту. Активність APX була визначена методом Бера — Ламберта при 290 нм.
Визначення активності протеази
Для визначення активності протеази 0,5 мл 1% легуміну (отриманому в 0,5 моль фосфатному буфері, pH 6) витримували з 0,2 мл ферментного екстракту при 45 °С протягом 1 години. Реакцію зупиняли додаванням 0,1 мл 40% трихлороцтової кислоти (ТСА). Активність протеази була розрахована на основі коефіцієнта екстинкції 21,5 ммоль-1см-1, а коефіцієнт абсорбції визначався при 280 нм.
Визначення малондіальдегіду (MDA)
Для визначення малондіальдегіду 0,2 г тканин свіжого листя подрібнювали з 5 мл 0,1% малондіальдегіду. Після центрифугування при 4000 об./хв протягом 20 хв, до супернатантів додавали 2,5 мл 0,5% тіобарбітурової кислоти (TBA) у 20% ТСА. Екстракти витримували на водяній бані при 95 °С протягом 30 хв та одразу ж охолоджували. Після цього всі зразки центрифугували при 4000 об./хв протягом 30 хв, а коефіцієнт абсорбції визначався при 532 нм та 600 нм. Концентрація MDA розраховували шляхом віднімання неспецифічної абсорбції при 600 нм від коефіцієнта абсорбції при 532 нм, використовуючи коефіцієнт згасання абсорбції 155 ммоль-1 см1.
Загальний вміст беталаїну
Екстракція беталаїну визначалася за допомогою модифікованого методу Френсіса. 5 г свіжого листя подрібнювали з 25 мл 20% етанолу та 0,5% розчину лимонної кислоти і залишали у темряві на 24 години. Після фільтрування супернатант використовували для очищення беталаїну методом звичайної колонкової хроматографії. У якості постійної фази використовували 10 г силікагелю 60 з сумішшю бінарного розчинника метанол/вода 8:2 об./об. з 1% об./об. безводної оцтової кислоти. Цей же бінарний розчинник застосовувався як елюація зі швидкістю потоку 0,7 мл/хв. Елюацію використовували для спектрофотометричного вимірювання коефіцієнта абсорбції при 535 нм.
Загальний вміст беталаїну визначали за допомогою наступного рівняння:
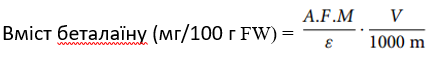
Де: A — абсорбція зразка;
M — середня молекулярна маса;
ε — молярний коефіцієнт поглинання;
V — об’єм екстракту;
m — маса рослинної твердої речовини, що використовується для вилучення;
DF — коефіцієнт розведення.
Статистичний аналіз
Статистичний аналіз виконувався за допомогою Однофакторного дисперсійного аналізу (версія 18; SPSS Inc., Чикаго, США). Середні значення стискання були представлені критерієм Данкана при P < 0,05.