Біохімія, що лежить в основі технології виробництва та механічної обробки насіння цукрових буряків (частина 2)

За матеріалами статті на тему «Біохімія, що лежить в основі технології виробництва та механічної обробки промислового насіння цукрових буряків» («The biochemistry underpinning industrial seed technology and mechanical processing of sugar beet») міжнародного журналу з біології рослин «Planta» (2019).
Вилучення та кількісне визначення абсцизової кислоти (АБК) та її метаболітів
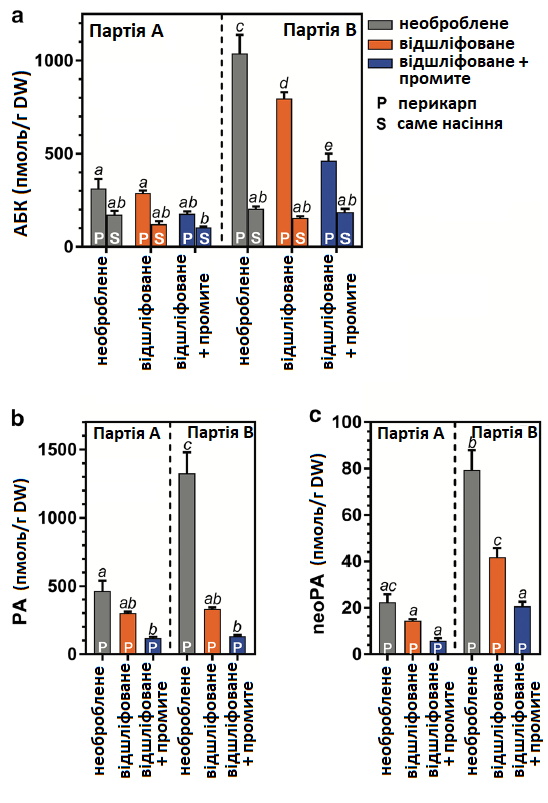
Щоб дослідити вміст фітогормону в перикарпі під час обробки, проаналізували зміни вмісту АБК та шлях деградації її метаболітів для окремих етапів обробки насіння. Аналіз проводили в ізольованому насінні та в відокремлених перикарпах (включаючи оперкулюми), отриманих із «необробленого», «відшліфованого» та «відшліфованого + промитого» насіння. Вміст АБК (Рис. 5а) у більшості зразків партії В був вищиv, ніж у партії А, при цьому вміст АБК у самому насінні завжди був нижчим, ніж у перикарпі. Вміст АБК в необробленому перикарпі партії В (1039 ± 101 пмоль/г DW) був у ~ 3,3 разів вищим, у порівнянні з необробленим перикарпом партії A (314 ± 52 пмоль/г DW). Унаслідок шліфування та промивання вміст AБК (а також (фазової кислоти) PA і (неофазової кислоти) neoPA) в перикарпах зменшився в ~ 2,3 разів у партії B (до 463 ± 39 пмоль AБК/г DW) та в ~ 1,8 разів у партії А (до 178 ± 13 пмоль AБК/г DW) (Рис. 5а).
Лише шліфування зменшило вміст АБК в перикарпі партії В в ~ 1,3 рази (до 796 ± 35 пмоль/г DW), але в партії А воно (шліфування) не мало суттєвого впливу. Вміст продуктів розпаду AБК — фазової кислоти (PA) і неофазової кислоти (neoPA) — (Рис. 5b, c) у перикарпах показав дуже подібні тенденції до АБК. У самому насінні вміст продуктів розпаду АБК був дуже низьким/нижчим межі виявлення (дані не вказані). Продукти розпаду демонструють, що відомі шляхи інактивації АБК є очевидними в тканині перикарпу обох партій. Висновок про те, що вміст АБК був більше ніж утричі вищим у перикарпах партії В, порівняно з партією А, підтверджує думку, що АБК сприяє більшій інгібуючій активності промивної води перикарпу партії В в аналізах проростання (Рис. 4). Вміст інших метаболітів АБК був або нижчим межі виявлення, або не показав істотних відмінностей між способами обробки насіння (дані не вказані).
Рис. 5. Визначення абсцизової кислоти (AБК) та її метаболітів — фазової кислоти (PA) та неофазової кислоти (neoPA) у насінні цукрових буряків. Ці метаболіти фітогормону були кількісно визначені в необробленому (сірий), відшліфованому (помаранчевий) і відшліфованому + промитому (синій) насінні партій А та В. Результати для самого насіння (S) та перикарпу (P) наводяться окремо. Вміст PA та neoPA був дуже низьким/нижчим межі виявлення у самому насінні, тому не вказується. Кожен стовпчик являє собою середнє значення ± SE 5 повторів. Кожен повтор складався з 50 насінин.
Осмоляльність, провідність та загальний вміст розчиненої речовини перикарпу
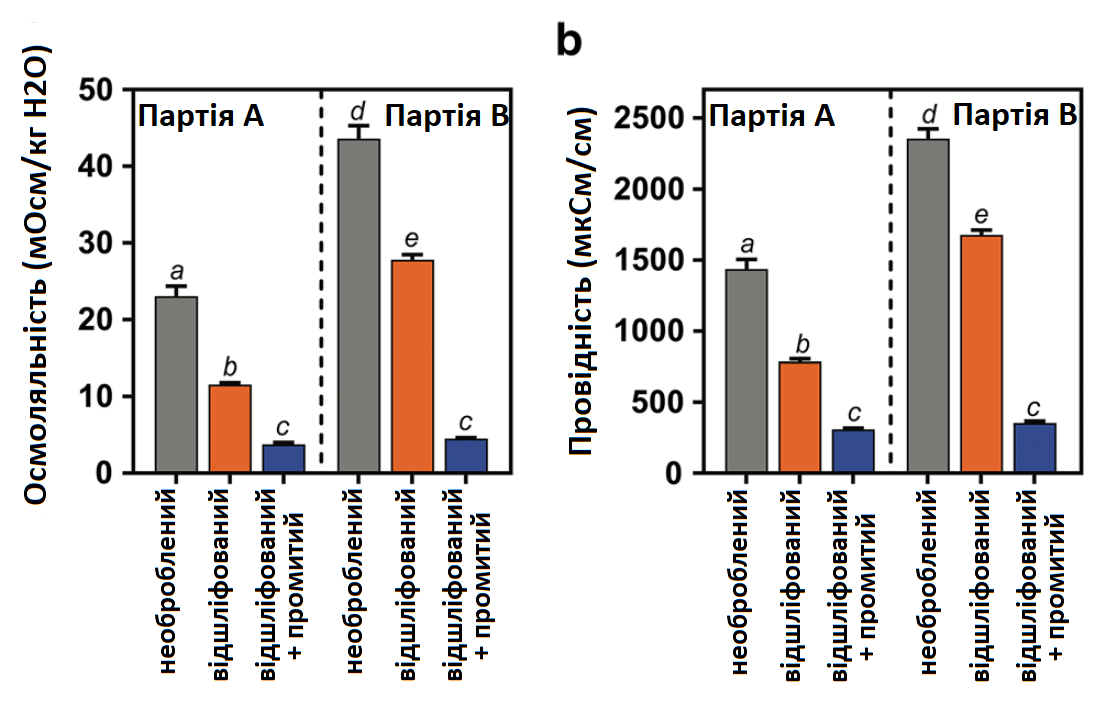
Для визначення вмісту іонів у порошкоподібних перикарпах необробленого, відшліфованого та відшліфованого + промитого насіння використовували воду в якості розчинника, вимірювали осмоляльність та провідність, які можуть використовуватися як приблизні значення для осмотично активних речовин, у тому числі солі. Спостережуване зменшення іонів внаслідок шліфування та промивання є подібним для обох партій насіння, але загальні значення вищі для партії В (Рис. 6а, b). Якщо порівняти вміст розчиненої речовини відшліфованого перикарпу з необробленим перикарпом, він мав у 2,0 рази нижчу осмолярність для партії А та в 1,6 разів нижчу осмоляльність для партії В, в 1,8 разів меншу провідність для партії А та в 1,4 разів нижчу провідність для партія B. Цікаво, що якщо порівнювати екстракти з відшліфованих + промитих перикарпів із екстрактами з необроблених перикарпів, вони мають в 6,1 разів нижчу осмолярність для партії А і в 9,7 разів нижчу осмоляльність для партії В, а також у 4,6 разів нижчу провідність для партії А і в 6,6 разів меншу провідність для партії В. Відмінності в осмоляльності між екстрактами перикарпа (необробленого проти відшліфованого + промитого порошкоподібного перикарпа в dH2O у співвідношенні 1:20) еквівалентні зміні концентрації NaCl від 13,1 до 3,1 ммоль для партії А і від 23,8 до 3,5 ммоль для партії В. Найбільше зменшення вмісту осмотично активних речовин у перикарпі було виявлено після застосування шліфування та промивання. Крім того, вища осмоляльність та провідність екстрактів перикарпу в партії В, порівняно з партією А, підтверджують думку, що вилучені сполуки можуть сприяти більш високій інгібуючій активності промивної води перикарпу в партії В, порівняно з партією А, в аналізах проростання (Рис. 4).
Рис. 6. a Осмоляльність та b провідність екстрактів промивної води перикарпу цукрового буряка є приблизними значеннями для солей, одержаних із необроблених (сірий), відшліфованих (оранжевий) та відшліфованих + промитих (синій) перикарпів партій А та В. Необроблене та відшліфоване насіння партії В мало вищі значення, ніж насіння партії А. Промивання відшліфованого насіння сильно зменшило значення обох партій. Кожна точка даних являє собою середнє значення ± SE 5 повторів. Кожен повтор являє собою екстракт промивної води з 50 насінин.
Скануюча електронна мікроскопія
Обробка також впливає на фізичні аспекти насіння. Шліфування має значний вплив на розмір та масу насіння (Рис. 1, Таблиця 1). Таким чином, використовувалася скануюча електронна мікроскопія (СЕМ), щоб отримати уявлення про морфологічні зміни в структурі перикарпу за рахунок обох етапів обробки насіння. Рис. 7a-c показує вид зверху поверхні та структурні деталі оперкулюму у необробленому, відшліфованому та відшліфованому + промитому насінні. Поверхневий вид необробленого насіння показує незмінену/природну структуру зовнішнього перикарпу (Рис. 7а). Вигляд поверхні відшліфованого насіння (Рис. 7b) дуже різниться, оскільки зовнішній шар перикарпу був значною мірою видалений, залишилися лише залишки великих клітин паренхіми на новоутвореній поверхні. Також видно (Рис. 7b), що всередині ешафоту клітин паренхіми є багато частинок різного розміру, які, швидше за все, є фрагментами клітинної стінки, які утворилися під час шліфування. Додатковий процес промивання (Рис. 7c) видаляє більшість цих фрагментів.
Спостереження в поперечному перерізі тканин перикарпу необробленого насіння (Рис. 7d, f, g, j) показують чітку відмінність між двома структурними шарами: внутрішнім шаром товстостінних клітин склеренхіми («внутрішній перикарп») і пористим зовнішнім шаром великих клітин паренхіми («зовнішній перикарп»). Ці два типи шарів можна розрізнити на основі структури клітин, розмірів, форми та товщини клітинних стінок. У деяких областях перикарпу відмінність між обома шарами дуже чітка (Рис. 7d), тоді як іноді можна виявити підшари клітин паренхіми (Рис. 7j), виходячи лише з розмірів клітин (більший на периферії, менший ближче до центру). При порівнянні поверхні внутрішнього і зовнішнього перикарпів, у зовнішньому перикарпі поперечний переріз проходить через внутрішню частину мертвих клітин паренхіми (виявляючи округлену внутрішню частину клітини), тоді як у внутрішньому перикарпі переріз проходить через апопласт (Рис. 7f, g, cf. h, i) (швидше за все, між клітинними стінками сусідніх клітин), залишаючи видимими клітинні з'єднання/плазмодесми (при великому збільшенні). Також було виявлено однакову організацію шарів в оперкулюмі та перикарпі.
Рис. 7. Морфологія поверхні та внутрішньої структури перикарпу насіння цукрового буряка за допомогою скануючої електронної мікроскопії (СЕМ). Зображення показують поверхню оперкулюму (a-c) або поверхню зламу вручну розламаних перикарпів (d-k) необробленого (UP) (a, d, f, g, j), відшліфованого (P) (b, e, h, i, k) та відшліфованого + промитого (PW) (c) насіння партії B. Масштабна шкала представляє 100 мкм кожного зображення.
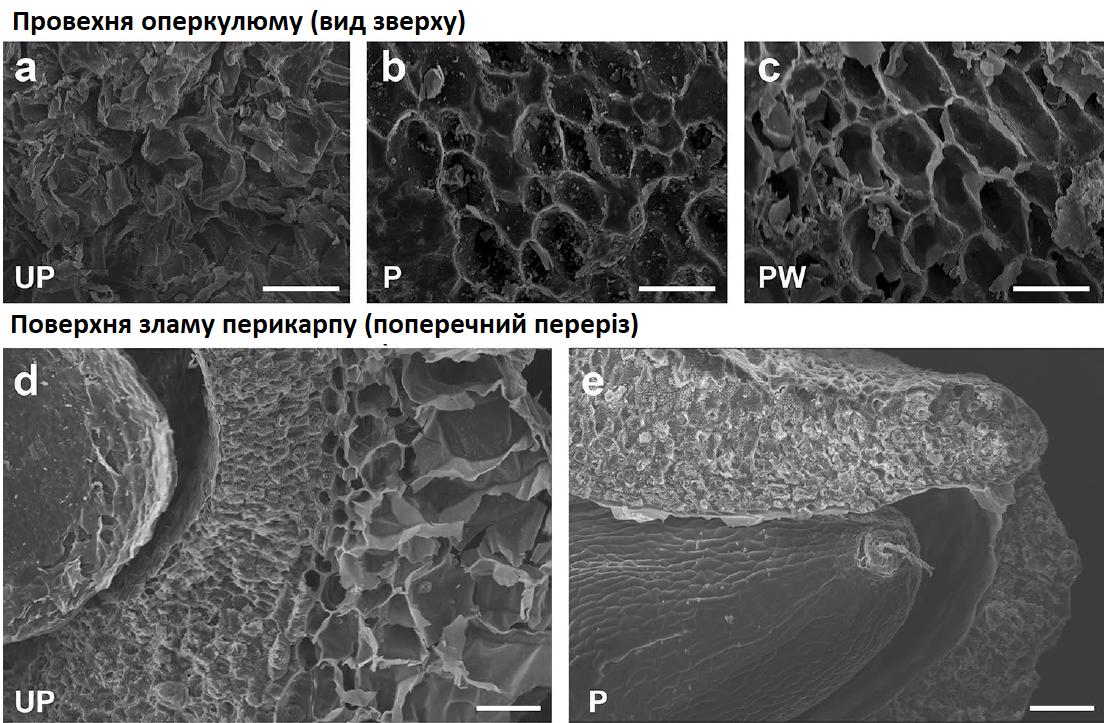
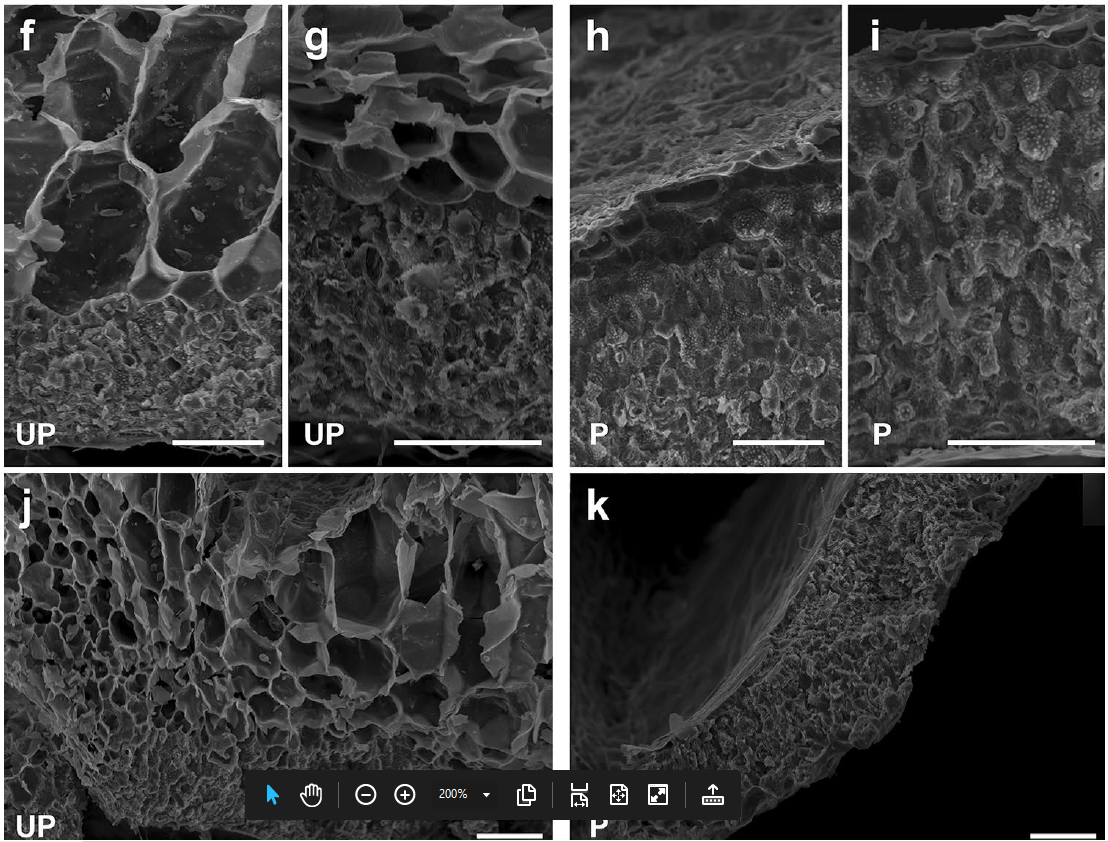
Морфологія відшліфованого насіння сильно змінюється, у порівнянні з необробленим насінням (Рис. 7te, k). Поверхня зламу вручну розламаних перикарпів показує, що шліфування часто видаляє всі, крім останнього або передостаннього ряду клітин у зовнішньому перикарпі. Шліфування не впливає на внутрішній перикарп (Рис. 7h, i). Менші клітини склеренхіми утворюють тверду і компактну структуру всередині перикарпу. Не було жодних доказів впливу промивання (без шліфування) на клітинні шари та структури в перикарпі та оперкулюмі (дані не наведені).
Обговорення
Перикарп насіння цукрового буряка як основний об’єкт шліфування та промивання у процесі промислової обробки для покращення якості та проростання насіння
Такі технології обробки, як шліфування та промивання, широко застосовуються для покращення фізіохімічних властивостей насіння, щоб забезпечити найякісніший продукт у ланцюжку постачання продуктів харчування. У сухому насінні B. vulgaris перикарп (насіннєва оболонка) є основним об’єктом шліфування та промивання. Метою шліфування в цілому є зменшення перикарпу, тим самим видалення інгібіторів проростання, гомогенізація насіння у фракцію робочого розміру та зміна геометрії (зменшення структури насінин, подібної до зірки), що забезпечує його дражування для використання у звичайних сівалках. Крім того, це покращує проростання для отримання рівномірних сходів та раннього розвитку молодих рослинок, щоб забезпечити потенціал врожайності за рахунок росту цукроносного коренеплоду (цукрового буряка). Незважаючи на це, структурні та текстурні зміни, а також основні біохімічні механізми, пов'язані з промисловими процесами шліфування та промивання, недостатньо досліджені. У деяких культурних рослин процес висушування насіння для покращення проростання змінює морфологію насіння або його оболонки лише в мінімальному ступені, у випадку B. vulgaris відбуваються більш значні морфологічні та біохімічні зміни, які необхідні для досягнення підвищеної фізіологічної якості та показників проростання насіння.
У даній статті показано, що промислове шліфування видалило від 31 до 35% зовнішньої тканини перикарпу насіння цукрового буряка і створило нову зовнішню поверхню з різними властивостями (Рис. 1, 7). Під час розвитку та сухого дозрівання насіння B. vulgaris формуються два основні шари тканини перикарпу: більш щільний внутрішній шар перикарпу, що складається з товстостінних клітин склеренхіми з безліччю чітко помітних плазмодесм (Рис. 7i), та більш пористий зовнішній шар перикарпу, що складається з великих, але тонкостінних клітин паренхіми (Рис. 7f). Відмінність між морфологічно різними внутрішнім та зовнішнім перикарпами бурякового насіння узгоджується з попередніми мікроскопічними дослідженнями. Тканини внутрішнього та зовнішнього перикарпів мають різні розміри та форми клітин, а також клітинні стінки. Обидва типи тканин присутні в нижній частині перикарпу і в оперкулюмі, який є ключовою особливістю насіння цукрового буряка, це куполоподібна або кришкоподібна структура, що покриває саме насіння з його закрученим зародком (Рис. 1). Оперкулюм і базальна пора обмежують поглинання кисню та води. Під час пізньої фази проростання цукрових буряків оперкулюм піднімається, а зародковий корінець виступає крізь щілину між нижнім перикарпом та оперкулюмом. Саме ізольоване насіння, з якого повністю видаляється перикарп, або насіння з видаленим перикарпом проростає швидше, в порівнянні з необробленим насінням, демонструючи те, що проростання цукрових буряків, принаймні частково, залежить від перикарпу.
Електронно-мікроскопічне дослідження показує, що шліфування видаляє більшість клітин паренхіми перикарпу, але немає жодних доказів того, що клітини склеренхіми були видалені (Рис. 7h, i). Тому можна припустити, що механічне видалення великих клітин паренхіми у зовнішньому перикарпі вимагає менших зусиль під час процесу шліфування, на відміну від клітин склеренхіми внутрішнього перикарпу. Як висновок, видалення зовнішнього пористого перикарпу досягається шліфуванням, що узгоджується з результатами інших досліджень. Однак, щоб виявити механічні відмінності між шарами паренхіми та склеренхіми їх реакція на шліфування вимагає більш детального біомеханічного дослідження перикарпу насіння цукрового буряка.
Шліфування створює нову штучну зовнішню поверхню насіння з різними властивостями та налипанням фракцій різного розміру всередині ешафоту клітин паренхіми (Рис. 7b). Промивання відшліфованого насіння цукрових буряків видаляє налиплі фракції (Рис. 7с), а також усуває необхідність розміщувати насіння оперкулюмом вниз у лунках для проростання. І шліфування, і промивання формують змінену нову структуру зовнішньої поверхні насіння, важливу для поліпшення адгезії покривних і дражувальних матеріалів. Тому важлива правильна інтеграція етапів шліфування та промивання у всю схему обробки насіння.
Шліфування та промивання сприяють видаленню хімічних інгібіторів проростання насіння цукрових буряків
У даній статті досліджувалося видалення шляхом шліфування та промивання двох груп хімічних сполук, локалізованих у перикарпі насіння цукрових буряків, що перешкоджають проростанню на прикладі насіннєвих партій А та В. Найважливішим висновком є те, що комбінована обробка «шліфування + промивання» призвела до значного покращення показників проростання обох партій насіння (Рис. 1) за рахунок комбінованого видалення механічних та інгібіторних обмежень. Відомо, що хімічні інгібітори цукрового буряка — це неорганічні іони, включаючи різні катіони (переважно Na+ та K+) та аніони (переважно Cl− та оксалат).
Відповідно до їхньої ролі як неорганічних інгібіторів, для пригнічення проростання насіння цукрових буряків були необхідні розчини NaCl у нормі щонайменше 100-300 ммоль (Рис. 4). Провідність промивної води перикарпу вважалася хорошим методом для моніторингу ефективності промислових процедур промивання. Було виявлено, що промивна вода перикарпу пригнічує проростання цукрових буряків, якщо значення провідності перевищувало ~ 10 мСм/см, що трохи менше, ніж 100 ммоль NaCl, використані у даних експериментах із пророщування (11,2 мСм/см; Рис. 4). Промивна вода перикарпу необробленого насіння цукрових буряків партій А і В мала значення провідності приблизно 1,9 мСм/см і 4,0 мСм/см відповідно, але це явно гальмувало проростання насіння обох партій (Рис. 4). Із цього можна зробити висновок, що солі у промивній воді перикарпу з партій А та В поодинці не можуть викликати інгібування проростання. Крім того, було виявлено явне зниження провідності під час шліфування та промивання щонайменше в 4,6 разів, що значно нижче концентрації NaCl, яка пригнічує проростання цукрових буряків, та підтверджує, що комбінована обробка «шліфування + промивання» зменшує вміст неорганічних солей у перикарпі до дуже низьких значень для покращення проростання насіння. Також було показано, що вимірювання осмоляльності промивної води перикарпу є відповідною альтернативою вимірювання провідності. Попередня робота також продемонструвала, що різноманітні фенольні сполуки, виділені або з перикарпу насіння червоного буряка, або з перикарпу насіння цукрового буряку, пригнічують проростання насіння та розвиток сходів таких цільових видів, як салат, крес-салат. Ці сполуки, які накопичуються в перикарпі, включаючи протокатехову, пара-гідроксибензойну, ванілінову, ферулову, р-кумарову та саліцилову кислоти, також були виявлені в подібних дослідженнях 2016 року. У сукупності «шліфування + промивання» ефективно зменшують вміст органічних та неорганічних сполук у перикарпі насіння цукрового буряка, але цих інгібіторів проростання недостатньо, щоб пояснити той факт, що промивна вода перикарпів насіння партії В мала більший гальмуючий вплив на проростання, у порівнянні з промивною водою перикарпів насіння партії А (Рис. 4).
Другою групою хімічних інгібіторів, що досліджувалися у перикарпі насіння цукрового буряка, були рослинні гормони — фітогормони. Попередні роботи продемонстрували, що тканина перикарпу містить високі рівні абсцизової кислоти (АБК) у мікромолярному діапазоні. Відомі вчені повідомляли про перевищення 1 мкмоль АБК на грам сухого перикарпу, а в даній статті це стосується необробленого насіння партії В, тоді як партія А мала нижчі значення ~ 0,3 мкмоль АБК на грам сухого перикарпу (Рис. 5а). Можна припустити, що більше ніж у 3 рази вищий вміст абсцизової кислоти (АБК) у партії В, порівняно з партією А, є основною причиною більшого інгібуючого впливу промивної води перикарпу насіння партії В, порівняно з партією А (Рис. 4). Шліфування та промивання ефективно зменшили вміст АБК в обох партіях насіння і були пов’язані з вимиванням абсцизової кислоти з перикарпу, в поєднанні з її біохімічною інактивацією за відомими 8 ′ та 9 ′ шляхами ферментативного гідроксилювання, що видно на прикладі метаболітів PA та neoPA (Рис. 5b, c). Цікаво, що стосовно загальної маси перикарпу, аналогічна кількість абсцизової кислоти була локалізована у внутрішньому перикарпі (53% — у партії В і 60% — у партії А), порівняно з зовнішнім перикарпом. Тому для зменшення вмісту абсцизової кислоти до низьких значень необхідна комбінована обробка «шліфування + промивання» насіння. 100 мкмоль АБК ефективно пригнічували проростання насіння партії В та А. На закінчення, у той час як різний вміст АБК у внутрішньому шарі перикарпу пояснює різницю між партіями В та А у швидкості проростання та інгібуючій активності промивної води перикарпу, комбіноване видалення різних груп інгібіторів (АБК, фенольних сполук, солей та інших) шляхом шліфування та промивання допомагає досягти повного потенціалу проростання кожної окремої насінини.
Отримання рівномірних сходів цукрових буряків залежить від якості процедур шліфування та промивання насіння
Шліфування та промивання насіння цукрових буряків збільшило відсоток нормальних сходів. Для ідеального застосування технології у насіннєвій галузі вирішальне значення має відсутність ненормальних або дуже низька поширеність ненормальних сходів. У 2015 році також повідомлялося про зменшення кількості ненормальних сходів після промивання насіння цукрового буряка, але на відміну від висновків даної статті, шліфування збільшило кількість ненормальних сходів, потенційно через механічні пошкодження зародкового корінця в ході самого шліфування. Це демонструє те, що оптимізація інженерії шліфувальної обробки має вирішальне значення для забезпечення рівномірності сходів. Нарешті, після шліфування та промивання проводиться процес праймінгу, покривання плівкою та дражування, що забезпечує найвищу якість насіння, готового до продажу.
Висновки
Проведене дослідження показало, що шліфування та промивання позитивно впливають на проростання насіння та розвиток молодих сходів; цей вплив стає сильнішим, якщо ці процеси поєднуються. Біохімічний механізм, що лежить в основі поліпшення якості насіння, включає суттєве зниження інгібіторів проростання, що знаходяться в перикарпі. Вміст абсцизової кислоти (АБК), її метаболітів та іонів у перикарпі значно знизився, особливо при поєднанні двох процесів обробки насіння. У процесі шліфування видаляється зовнішня тканина перикапру (паренхіма), тоді як щільніша тканина (склеренхіма) залишається неушкодженою. А поєднання процесів шліфування та промивання є корисним інструментом для покращення якості та проростання промислового насіння.